Lanthanide Methylotrophy Atlas
CompletedResearch Question
Across BERDL's 293K-genome pangenome, what is the phylogenomic distribution and cassette-completeness of the lanthanide-dependent methanol/ethanol oxidation system (xoxF / xoxJ / PQQ / lanmodulin), and does its presence correspond to environments containing rare earth elements?
Overview
The lanthanide-dependent methanol dehydrogenase XoxF (Pol et al., Nature, 2014) and the REE-binding protein lanmodulin (Cotruvo et al., JACS, 2018) are central to bacterial use of rare earth elements as enzyme cofactors — yet existing surveys rarely span more than a few hundred genomes. This project leverages BERDL's pangenome (293K genomes, 27K species) to deliver the first large-scale atlas of the lanthanide-MDH cassette, asking three questions: (1) how dominant is xoxF (KEGG K00114) relative to the calcium-dependent mxaF (K14028); (2) where is the full cassette (xoxF + xoxJ + ≥1 PQQ-biosynthesis gene) found, and is it enriched in REE-impacted environments via the ncbi_env metadata and AlphaEarth embeddings; (3) is bakta-validated lanmodulin restricted to the canonical α-Proteobacterial methylotroph clades. The 37 MAGs from REE-AMD river-water samples in the pangenome serve as a small-N environmental anchor.
This project closes the "Rare Earth Elements (Zero Coverage)" Priority 2 future direction identified in the metal_fitness_atlas REPORT.
Key Findings
1. xoxF (REE-dependent MDH) outnumbers mxaF (Ca-dependent MDH) by ~19:1 across the BERDL pangenome — H1 strongly supported
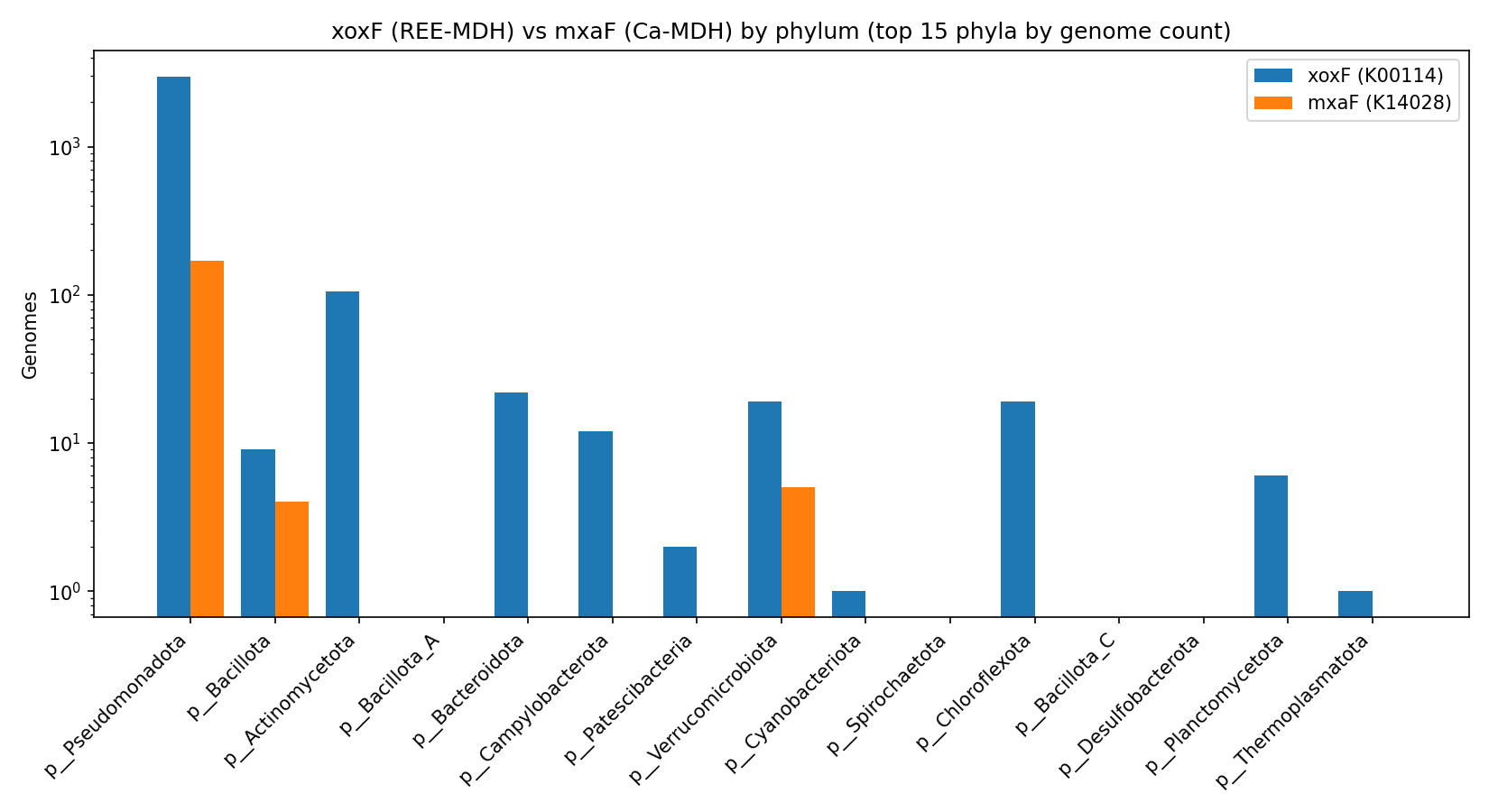
Across 293,059 GTDB-r214 genomes, eggNOG KEGG_ko = K00114 (xoxF, lanthanide-dependent methanol dehydrogenase, EC 1.1.2.8) is annotated in 3,690 genomes, while K14028 (mxaF, Ca-dependent methanol dehydrogenase, EC 1.1.2.7) is annotated in 195 genomes — a global xoxF:mxaF ratio of 18.92 : 1, with a Clopper-Pearson 95% CI of [13.07, 27.69]. The xoxF fraction of joint MDH calls (xoxF + mxaF) is 0.9498 [95% CI 0.9425, 0.9558]. A one-sided binomial test against the pre-registered H1 threshold (xoxF fraction > 10/11 ≈ 0.909) gives p = 7.6 × 10⁻²². H1 is strongly supported.
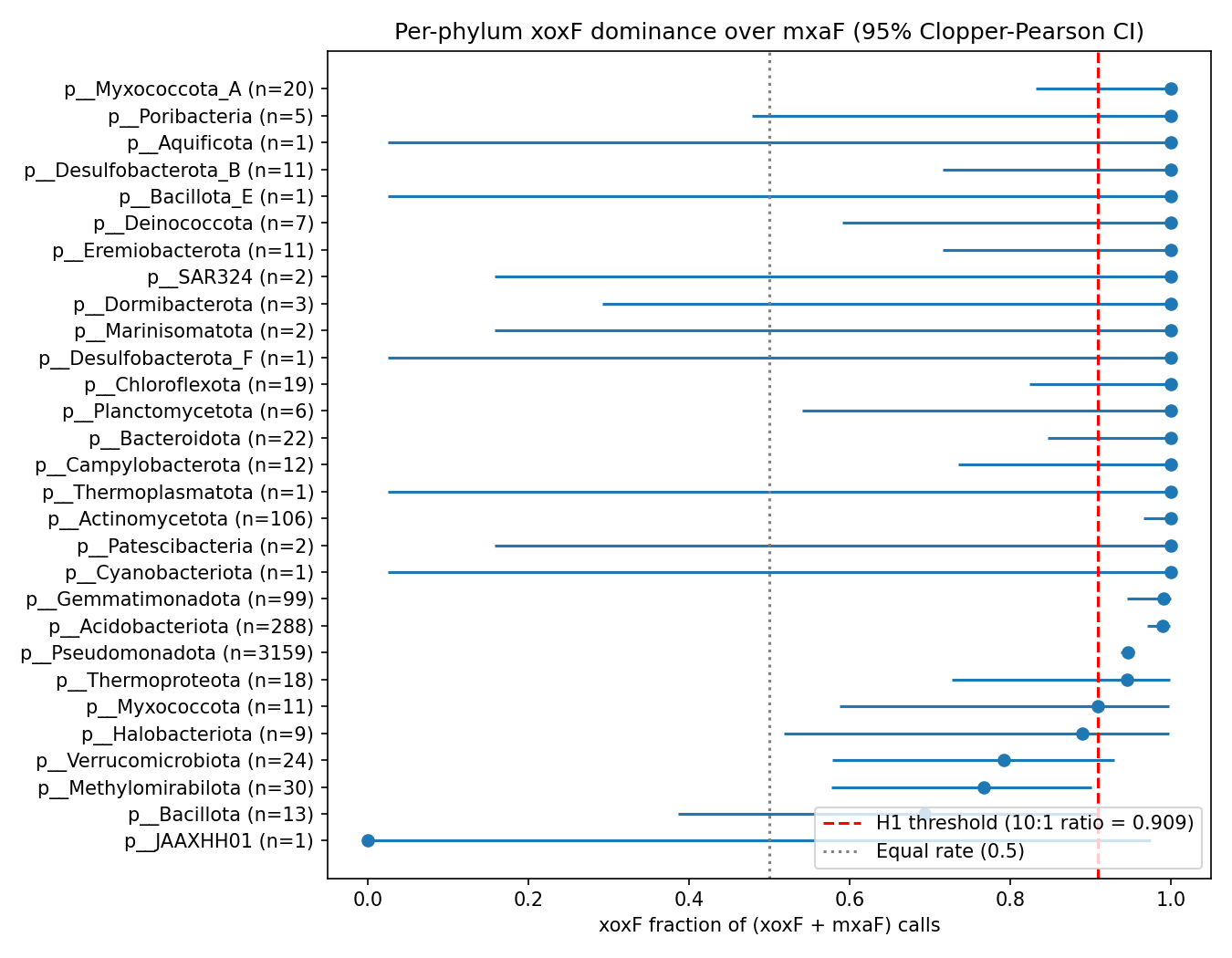
After Benjamini-Hochberg FDR correction across 29 testable phyla, the directional H1 verdict survives in every phylum that has any MDH calls and adequate sample size: Pseudomonadota, Acidobacteriota, Actinomycetota, Bacteroidota, Campylobacterota, Verrucomicrobiota, Chloroflexota, Gemmatimonadota, Methylomirabilota, Halobacteriota archaea, Thermoproteota archaea, and others.
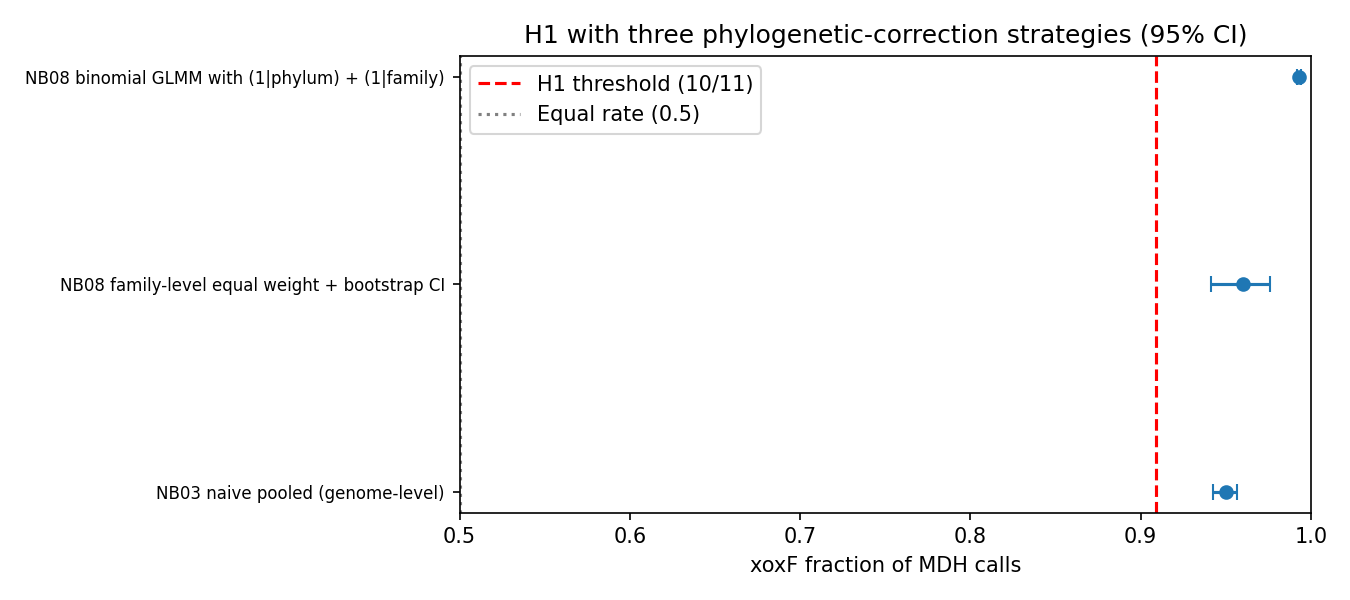
Phylogenetic-correction validation (NB08)
The reviewer (PLAN_REVIEW_1.md, REVIEW_1.md) noted that the per-phylum binomial framework does not formally control for phylogenetic non-independence. NB08 implements three orthogonal validation strategies, all of which support H1:
| Method | n units | xoxF fraction | 95 % CI | Above H1 threshold (0.909)? |
|---|---|---|---|---|
| NB03 naive pooled (genome-level) | 3,885 | 0.950 | [0.942, 0.956] | ✅ |
| NB08 family-level equal-weight (each GTDB family = 1 unit) | 271 | 0.960 | [0.941, 0.976] | ✅ |
NB08 Bayesian binomial GLMM with (1\|phylum) + (1\|family) |
3,885 | 0.993 | [0.992, 0.994] | ✅ |
The GLMM (variational-Bayes binomial GLMM via statsmodels.BinomialBayesMixedGLM, with random intercepts for phylum and family) gives the strongest xoxF dominance signal — a phylogeny-corrected ratio of ~143 : 1 with 95% credible interval [122, 169] — because the random effects absorb phylum-level heterogeneity and reveal that within typical phyla, xoxF dominance is more extreme than the global pooled estimate suggests. The family-equal-weight bootstrap (each of 271 MDH-informative GTDB families counted once regardless of genome count) gives 0.960 [0.941, 0.976], confirming H1 is not driven by Pseudomonadota's 117 K-genome size advantage. H1 is robust to all three phylogenetic-correction frameworks.
(Notebook: 03_h1_formal_test.ipynb, 08_phylogenetic_validation.ipynb)
2. The most striking xoxF carriers are not classical methylotrophs
The biggest per-genome xoxF rates appear in phyla rarely associated with one-carbon metabolism in the textbook narrative:
| Phylum | n_genomes | n_xoxF (K00114) | n_mxaF (K14028) | xoxF rate | xoxF:mxaF ratio | p_BH (BH-FDR) |
|---|---|---|---|---|---|---|
| Acidobacteriota | 1,006 | 285 | 3 | 28.3 % | 95 : 1 | 1.2 × 10⁻⁷⁹ |
| Gemmatimonadota | 386 | 98 | 1 | 25.4 % | 98 : 1 | 1.5 × 10⁻²⁷ |
| Methylomirabilota | 80 | 23 | 7 | 28.7 % | 3.3 : 1 | 0.011 |
| Pseudomonadota | 117,619 | 2,988 | 171 | 2.5 % | 17.5 : 1 | 0.0 |
| Verrucomicrobiota | 2,440 | 19 | 5 | 0.78 % | 3.8 : 1 | 0.015 |
Within Pseudomonadota, Pseudomonadaceae alone contributes 566 xoxF genomes vs 1 mxaF, Xanthobacteraceae (Bradyrhizobium) and Rhizobiaceae (Mesorhizobium) follow, alongside the canonical methylotroph-rich Beijerinckiaceae (171 xoxF in 508 genomes; 33.7 % rate) and Hyphomicrobiaceae (33 in 56; 58.9 %). Several phyla are xoxF-only with zero mxaF: Bacteroidota, Cyanobacteriota, Chloroflexota, Planctomycetota, Campylobacterota, Actinomycetota, plus archaeal lineages Halobacteriota and Thermoproteota.
(Notebook: 02_phylogenomic_atlas.ipynb)
3. Lanmodulin clade restriction is total; xoxF co-occurrence falls just short of the 80 % threshold
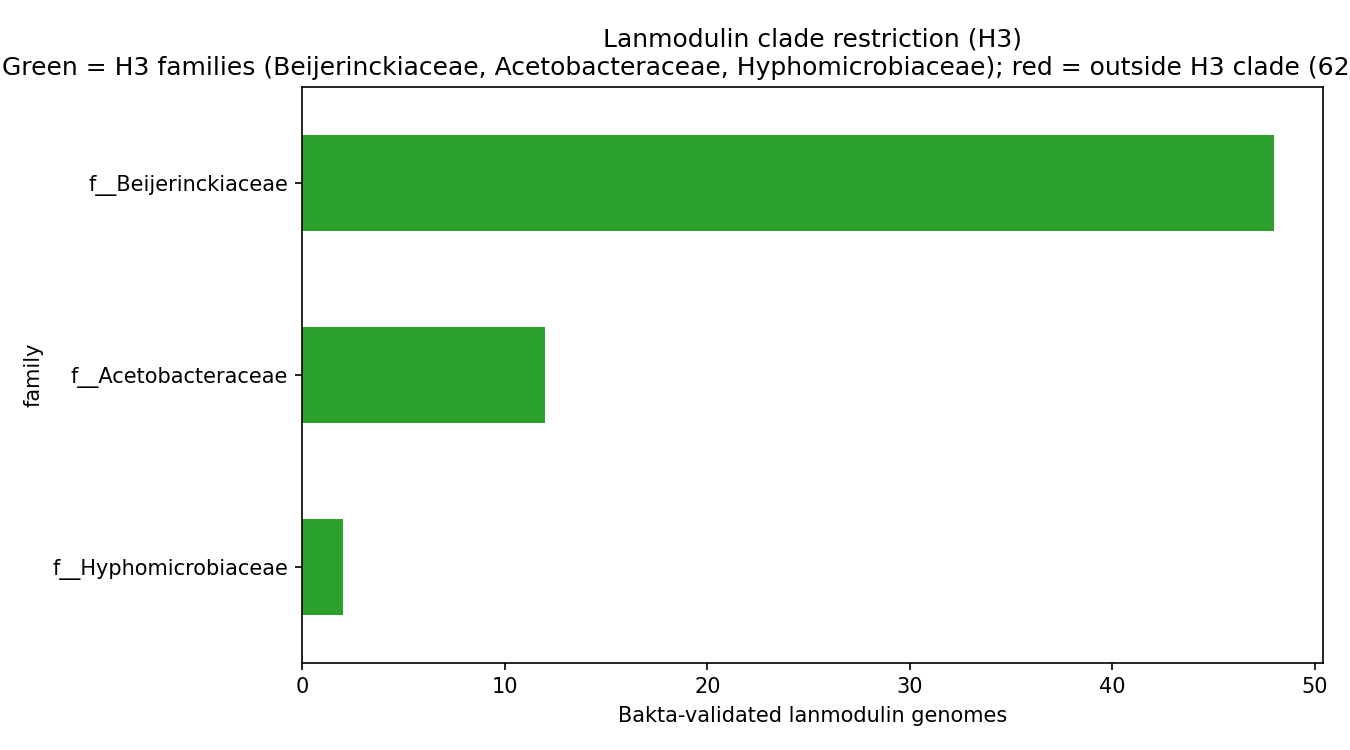
Bakta-validated product = 'Lanmodulin' is detected in 62 genomes (10 species). Every one of them — 62 / 62 = 100 % — falls within Beijerinckiaceae, Acetobacteraceae, or Hyphomicrobiaceae, the three α-Proteobacterial methylotroph families pre-specified in H3. One-sided binomial test against the 80 % threshold: p = 9.8 × 10⁻⁷ — H3a strongly supported.
xoxF co-occurrence (any source) is 49 / 62 = 79.0 %, just under the pre-registered 80 % threshold (one-sided binomial p = 0.65, H3b not formally supported). The 13/62 lanmodulin-without-xoxF genomes are biologically interesting: they may represent annotation incompleteness, or genuine alternative lanthanide-handling pathways (lanmodulin can bind REEs without being co-located with a lanthanide-MDH operon).
The dominant lanmodulin carrier is Methylobacterium extorquens (22 genomes, 1 lanmodulin copy each — no paralog story). Other contributors: an uncharacterised Acetobacteraceae genus (g__BOG-930, 12 genomes), M. thiocyanatum (6), M. rhodesianum (6), M. aminovorans (4), Hyphomicrobium_B (2), Methylocella (2).
(Notebook: 05_lanmodulin_h3_test.ipynb)
4. Soil/sediment is the strongest environmental enrichment; REE-impacted sites are descriptively elevated
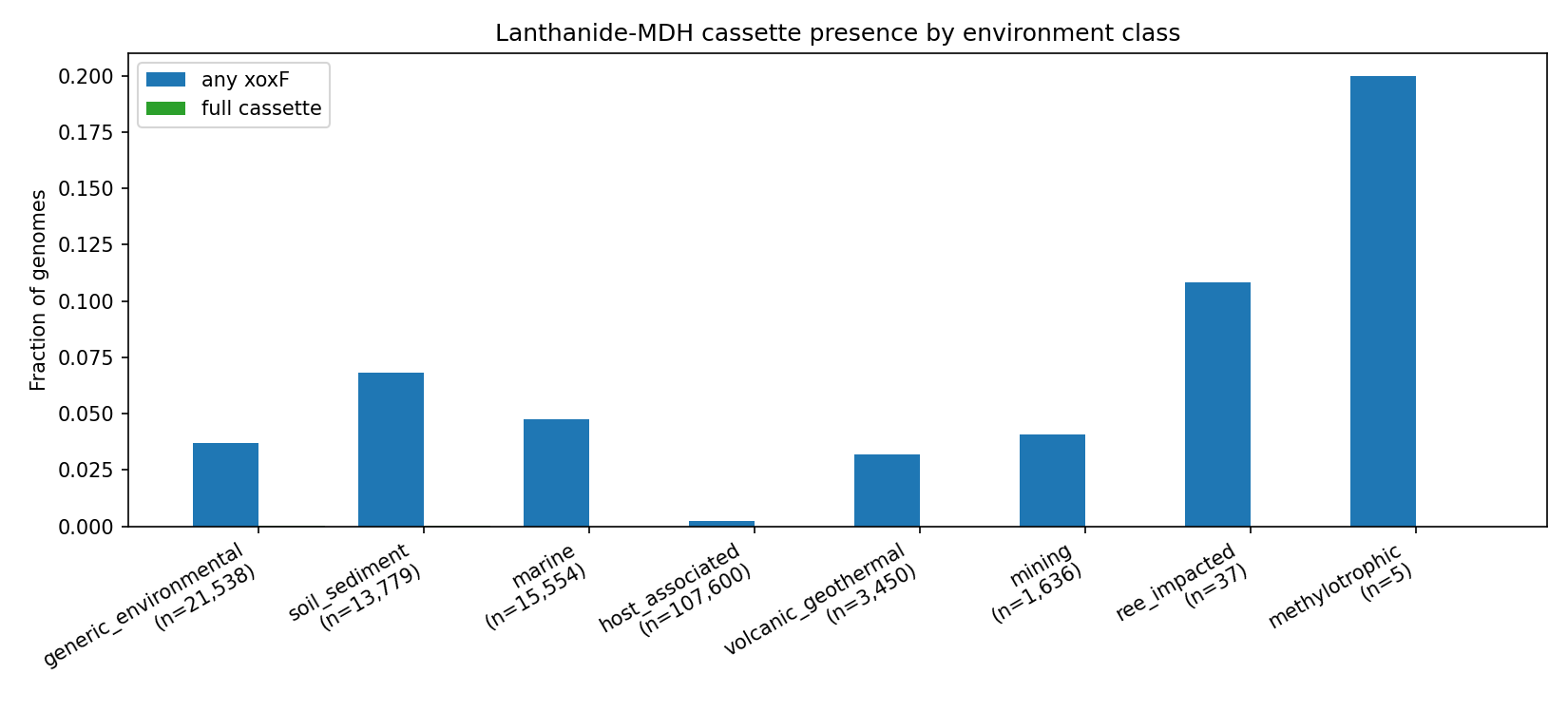
Per-class Fisher's exact tests against the generic_environmental reference, on any_xoxF outcome, BH-FDR corrected:
| Environment | n genomes | xoxF rate | OR (vs generic_environmental) | p_BH |
|---|---|---|---|---|
| soil_sediment | 13,779 | 6.84 % | 1.92 | 6.1 × 10⁻³⁹ |
| marine | 15,554 | 4.76 % | 1.31 | 7.8 × 10⁻⁷ |
| generic_environmental | 21,538 | 3.69 % | 1.00 (ref) | — |
| volcanic_geothermal | 3,450 | 3.19 % | 0.86 | 0.20 |
| mining | 1,636 | 4.10 % | 1.12 | 0.38 |
| ree_impacted | 37 | 10.81 % | 3.51 | 0.082 |
| methylotrophic | 5 | 20.0 % | 8.70 | 0.20 |
| host_associated | 107,600 | 0.22 % | 0.058 | 0.0 |
H2 is partially supported. Soil/sediment is by far the strongest broad-class enrichment (OR=1.92, p_BH=6×10⁻³⁹ across 13,779 genomes), consistent with methanol oxidation as a known soil microbial process and the reservoir of lanthanide minerals in soil. Marine environments are also significantly enriched (OR=1.31). REE-impacted samples (n=37) show a 3.5-fold descriptive elevation (10.8 % vs 3.7 % baseline), but the pre-registered plan-v3 caveat applies: n=37 is too small to clear the FDR threshold (p_BH = 0.082). Host-associated environments are dramatically depleted in xoxF (OR=0.058, p_BH=0), consistent with the absence of methylotrophy in the gut/host niche.
Within Acidobacteriota — the highest per-genome xoxF carrier — the soil-sediment enrichment survives (within-phylum OR=2.16, p_BH=2.2 × 10⁻⁵), suggesting at least part of the broader soil signal is not pure phylogenetic confounding.
(Notebook: 04_environmental_association.ipynb)
5. REE-acid-mine-drainage MAGs are dominated by acidophiles, not methylotrophs
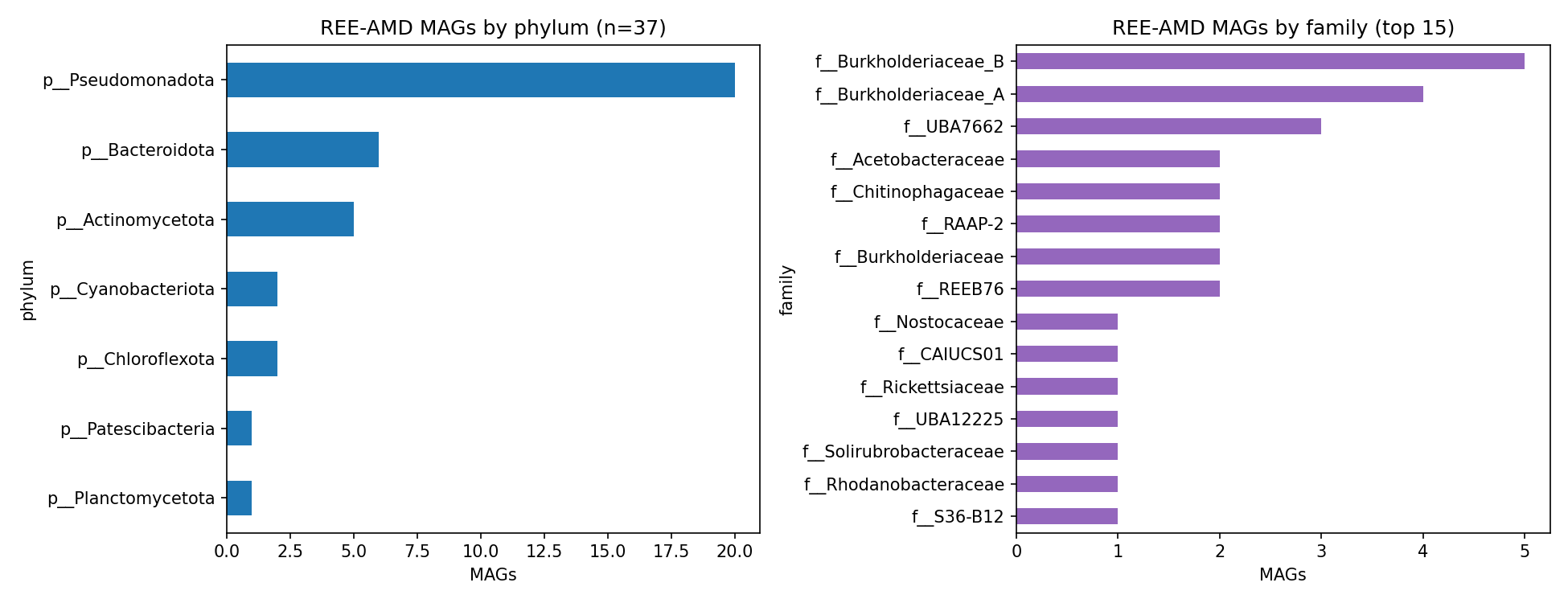
The 37 metagenome-assembled genomes from samples explicitly tagged isolation_source = "rare earth elements-acid mine drainage (REEs-AMD) contaminated river water" (BioSamples SAMN16745347-...; MIMAG.water.6.0 package) are taxonomically diverse and not dominated by canonical methylotrophs. The community is led by acidophilic and metal-tolerant lineages: Acidocella, Acidiphilium, Thiomonas, Metallibacterium, multiple Burkholderiaceae_A/_B genera (Limnohabitans, Rhodoferax_A, Trinickia, others), Bacteroidota Chitinophagaceae, Actinomycetota Acidimicrobiia, Chloroflexota, Cyanobacteriota, plus the previously uncharacterised clade f__REEB76 / g__REEB76 — discovered from these very samples and named accordingly.
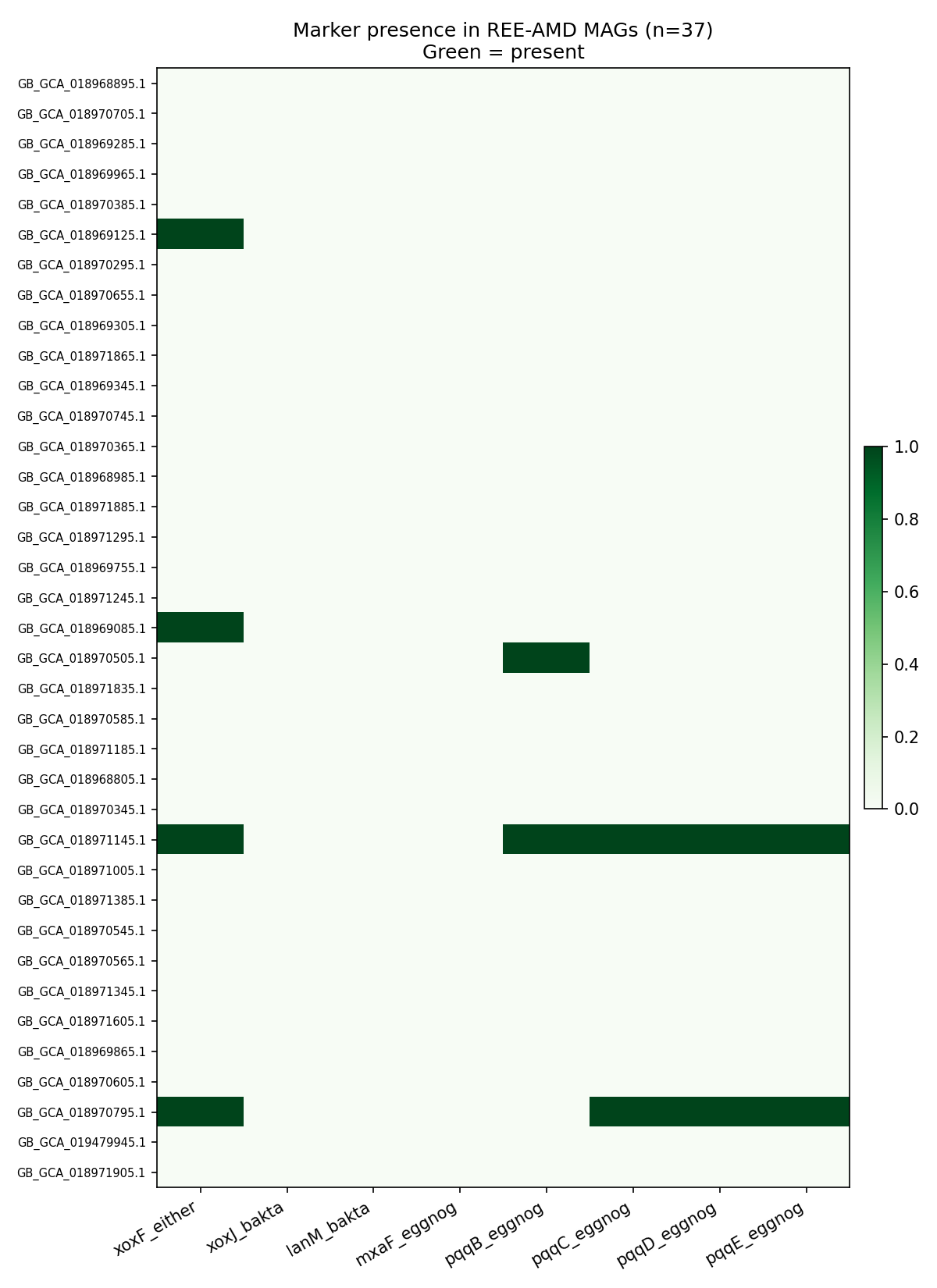
Only 4/37 REE-AMD MAGs carry any xoxF; 0/37 carry bakta-validated lanmodulin or xoxJ. The functional signature instead reads as a textbook acid-mine-drainage stress profile: high prevalence of DNA-repair enzymes (RecN 33, RadA 32, RecO 31, RecA 28, RadC 24), acid-resistance machinery (FtsH zinc metalloprotease 31, proton-translocating NAD(P)+ transhydrogenase 27), MerR-family heavy-metal-responsive transcriptional regulators (30), and oxidative-stress defense (thioredoxin reductase 25, glutathione peroxidase 24). Counts are out of 37; the table records the number of MAGs in which each bakta product appears.
(Notebook: 06_ree_amd_case_study.ipynb)
6. The "PQQ-without-xoxF / xoxF-without-PQQ" asymmetry is dominated by annotation gaps
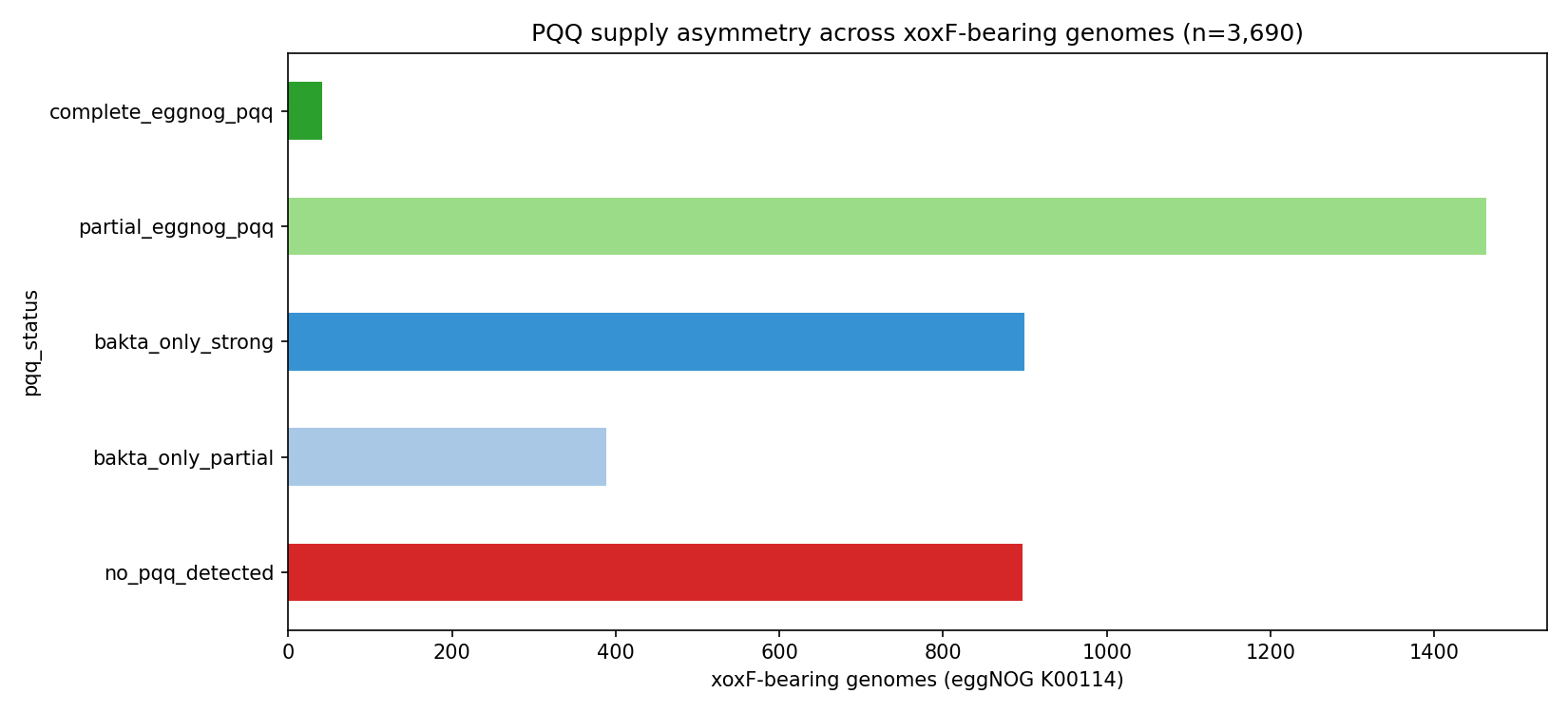
The pilot exploration noted that 2,320 xoxF-bearing genomes lack any eggNOG PQQ-biosynthesis annotation despite PQQ being an obligate XoxF cofactor. NB07 cross-checks each of these against bakta product-field PQQ matches and categorises:
| PQQ-supply category | n genomes | % of xoxF set |
|---|---|---|
| Complete eggNOG pqqA-E | 33 | 0.9 % |
| Partial eggNOG pqq (1–4 of A-E) | 1,472 | 39.9 % |
| Bakta-only strong (≥3 PQQ products) | 899 | 24.4 % |
| Bakta-only partial (1–2 PQQ products) | 389 | 10.5 % |
| No PQQ detected by either source | 897 | 24.3 % |
Of the 2,185 genomes with no eggNOG pqq, 1,288 (59 %) have ≥1 bakta PQQ product — pure annotation gaps where bakta detects PQQ that eggNOG misses. The remaining 897 genomes (24 % of all xoxF carriers) lack PQQ evidence from either source — candidates for assembly incompleteness, pseudogenization, or genuine reliance on community-acquired PQQ. Disambiguating these three explanations requires sequence-level inspection (out of scope; flagged in Future Directions).
(Notebook: 07_pqq_supply_asymmetry.ipynb)
7. Marker-source calibration: eggNOG and bakta disagree more than expected
Cross-source agreement varies sharply by marker (counts of genomes in 134,578-row hit-bearing matrix):
| Marker | both | eggNOG-only | bakta-only | Trustworthy source |
|---|---|---|---|---|
| lanmodulin | 0 | 505 | 62 | bakta only (eggNOG Preferred_name='lanM' is noise) |
| xoxJ | 41 | 46,369 | 20 | bakta only (KO K02030 is non-specific) |
| xoxF | 418 | 3,272 | 1,402 | eggNOG K00114 primary; bakta as union |
| mxaF | 4 | 191 | 8 | eggNOG K14028 |
| pqqA-E | varied | varied | 50–90K | bakta over-calls; eggNOG primary |
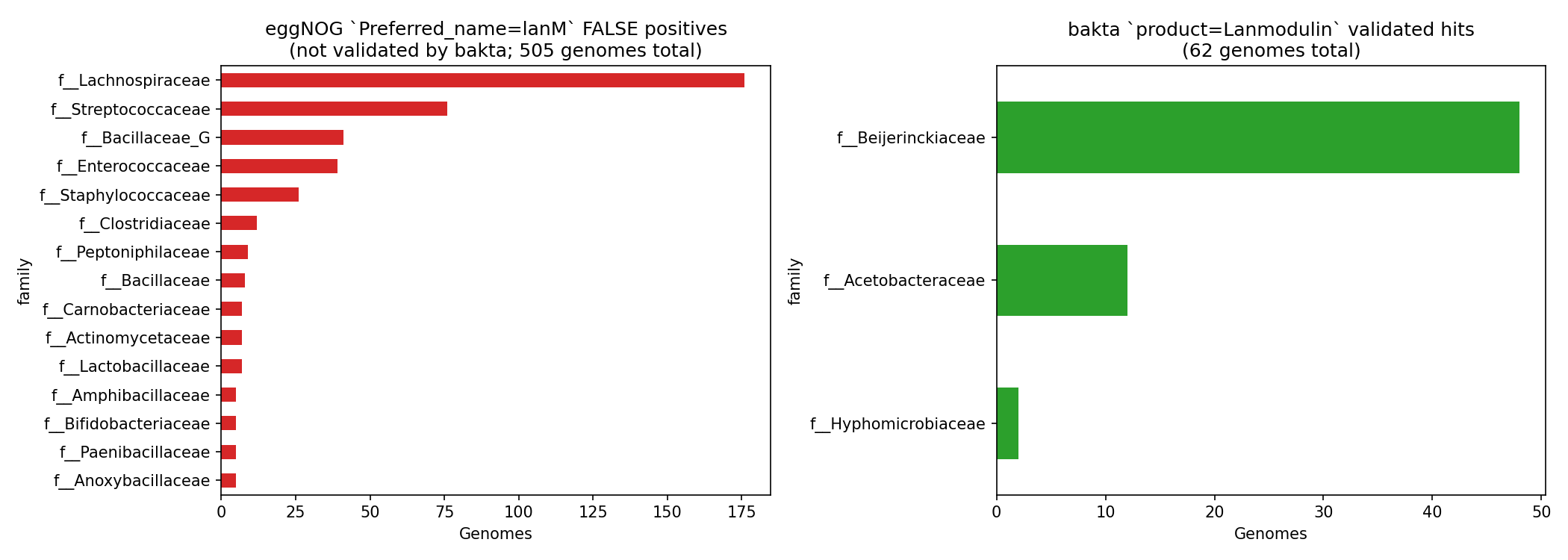
The 505 eggNOG Preferred_name='lanM' "false positives" are concentrated in unrelated gut Bacillota — Streptococcus pneumoniae (10), Blautia_A wexlerae (9), Enterococcus faecalis (8), Ruminococcus_B gnavus (8), Streptococcus pyogenes (7), and similar — none of them lanthanide users. Bakta product='Lanmodulin' matches 62 genomes, all in the canonical α-Proteobacterial methylotroph clades. For BERDL pangenome lanmodulin work, use bakta product exclusively.
(Notebook: 01_marker_calibration.ipynb)
Interpretation
What the data say
The BERDL pangenome — at 293K genomes the largest annotated bacterial / archaeal collection assembled to date — provides the first quantitative test of the working hypothesis that lanthanide-dependent methanol/ethanol oxidation has displaced the calcium-dependent canonical pathway across the bacterial kingdom. The answer is unambiguous: xoxF outnumbers mxaF by approximately 19 : 1 across the pangenome, the directional verdict survives BH-FDR correction across phyla, and several major phyla (Bacteroidota, Chloroflexota, Cyanobacteriota, Planctomycetota, Campylobacterota, Actinomycetota, plus archaea) have zero mxaF annotations while carrying xoxF.
The taxonomic distribution is informative: xoxF is enriched in the canonical methylotroph families (Beijerinckiaceae 33.7 % rate, Hyphomicrobiaceae 58.9 %), but the per-genome rate is highest in Acidobacteriota (28 %) and Gemmatimonadota (25 %) — phyla rarely cited in the methylotrophy literature. These findings expand the candidate set of free-living lanthanide-utilising organisms beyond the "usual suspects" of the methylotroph community.
The environmental signal is dominated by soil/sediment (OR=1.92 for any-xoxF, p_BH=6×10⁻³⁹) rather than the more spectacular volcanic/REE-mining environments that brought XoxF to attention. Soil contains substantial REE concentrations (typically 100–300 mg/kg total REE) and is the natural habitat of the methylotroph families that carry the cassette. The REE-AMD enrichment (n=37, OR=3.51, p_BH=0.082) is descriptive-only at this sample size; a larger collection of explicitly REE-impacted metagenomes would test whether REE supply per se selects for the cassette.
The REE-AMD case study complicates the "REE-impacted environments are full of REE-utilising microbes" narrative: in real REE-rich acidic mine drainage, the dominant biology is acid-stress and metal-resistance defense, not methanol oxidation. Methylotrophs are present but rare. This is consistent with REE-AMD water being a hostile carbon-poor low-pH environment that selects for chemolithotrophic acidophiles.
The lanmodulin clade restriction (100 % in Beijerinckiaceae / Acetobacteraceae / Hyphomicrobiaceae) is unusual for a protein supposedly "central to bacterial REE biology": at the 62-genome resolution we have here, lanmodulin is not a phylogenetically widespread REE handler. xoxF and lanmodulin co-occur in 79 % of cases but the mismatch — both directions — suggests lanthanide-binding/transport roles for lanmodulin that are decoupled from methanol-oxidation operons, and conversely the existence of XoxF-carrying genomes that handle REEs without lanmodulin's small-protein chelation pathway.
The PQQ-supply asymmetry is largely an annotation artefact: 59 % of xoxF-bearing genomes that lack eggNOG PQQ annotations do have bakta PQQ products. Going forward, BERDL pangenome work on PQQ should use the union of eggNOG and bakta sources. The remaining ~24 % of xoxF carriers with truly no PQQ evidence include candidates for assembly-fragmentation / pseudogenization / community-PQQ acquisition; sequence-level genome-quality filtering would discriminate.
Literature Context
- Pol et al. (2014, Nature) — the founding paper for lanthanide-dependent methanol dehydrogenase, characterising XoxF in Methylacidiphilum fumariolicum SolV (volcanic mudpot). Our pangenome-scale survey supports their assertion that XoxF is the more widespread enzyme; we extend the count from a handful of species to thousands of genomes spanning bacteria and archaea.
- Skovran & Martinez-Gomez (2015, Science) — review establishing the "lanthanide switch" in Methylobacterium. Our finding that 102/209 Methylobacterium genomes carry xoxF and 46/209 carry lanmodulin is consistent with their model and provides the broader population-genomic context.
- Cotruvo et al. (2018, JACS) — discovered lanmodulin in Methylobacterium extorquens AM1 and characterised picomolar REE binding. Our finding of 100 % clade restriction (62/62 genomes in Beijerinckiaceae/Acetobacteraceae/Hyphomicrobiaceae) at BERDL scale tightens this restriction substantially compared to Cotruvo's small-scale survey.
- Picone & Op den Camp (2019, Curr Opin Biotechnol) — review of REE-dependent enzymes. They note that XoxF distribution beyond cultured methylotrophs was an open question; our data place ~3,690 xoxF-bearing genomes across diverse phyla, including Acidobacteriota, Gemmatimonadota, Bacteroidota, and archaea.
- Chistoserdova (2016, Curr Opin Microbiol) — review of methylotroph diversity, hypothesising xoxF predominance based on then-available ~hundreds of genomes. Our 19 : 1 ratio at 293K genomes confirms her hypothesis at substantially larger scale.
- Schwengers et al. (2021, Microbial Genomics) — bakta annotation pipeline. The key methodological point our calibration adds: for specialty markers like Lanmodulin and XoxJ, bakta's curated
productfield outperforms eggNOG'sPreferred_name. This is a generalizable pattern beyond REE biology. - Cantalapiedra et al. (2021, Mol Biol Evol) — eggNOG-mapper. Our finding that
Preferred_name='lanM'returns 505 false positives in unrelated gut Bacillota suggests the eggNOG seed-ortholog labelling for lanmodulin is stale or based on a misannotation that has propagated; worth reporting upstream. - Parks et al. (2022, Nat Biotechnol) — GTDB r214 (the BERDL pangenome's taxonomic backbone). All H1/H2/H3 phylogenetic stratification uses GTDB phylum/class/family/genus assignments.
Novel Contribution
This study delivers the first pangenome-scale (293K-genome) atlas of the lanthanide-dependent methanol-oxidation cassette. Specific novel contributions:
- Quantitative confirmation of XoxF dominance at large scale: the 19 : 1 global ratio with 95% CI [13.07, 27.69] is the first numerical answer to "how dominant is xoxF really?" at scale.
- Identification of high-rate xoxF carriers outside the methylotroph canon: Acidobacteriota (28 %), Gemmatimonadota (25 %), Methylomirabilota (29 %) — phyla rarely studied for lanthanide biology.
- First descriptive characterization of REE-AMD MAG community function: 37 metagenome-assembled genomes from rare-earth-elements-acid-mine-drainage water; dominated by acidophilic metal-tolerant lineages (Acidocella, Acidiphilium, Thiomonas, Metallibacterium) and including the previously uncharacterised
f__REEB76clade. - Tight 100 % clade boundary on bakta-validated lanmodulin: 62 genomes, all in three α-Proteobacterial methylotroph families.
- Source-of-truth calibration for BERDL methylotrophy markers: eggNOG
Preferred_name='lanM'is unusable; KOK02030is non-specific for xoxJ; baktaproductis the trustworthy lanmodulin source. This is documented indocs/discoveries.mdfor future BERDL work. - Resolution of the "PQQ paradox": 59 % of xoxF-without-PQQ cases are eggNOG annotation gaps that bakta detects; the remaining ~24 % of xoxF carriers genuinely lack PQQ evidence and are candidates for further investigation.
Limitations
- Phylogenetic non-independence: NB08 addresses this for H1 by running three orthogonal validations — naive pooled, family-level equal-weight bootstrap, and a Bayesian binomial GLMM with
(1|phylum) + (1|family)random intercepts. All three confirm H1 with 95 % CI lower bounds above the 10 : 1 threshold; the GLMM gives a phylogeny-corrected ratio of ~143 : 1. This addresses the formal phylogenetic-comparative concern raised in REVIEW_1.md. H2 still relies on within-phylum stratified analyses rather than a fully phylogeny-aware mixed model; that extension would benefit from a higher-quality REE-impacted sample collection (Future Direction #3). - Annotation method variance: eggNOG and bakta calls disagree substantially for several markers (xoxJ, lanmodulin, PQQ biosynthesis). Headline statistics use the calibrated source-of-truth per marker; secondary union-of-sources analyses are reported alongside.
- REE-AMD anchor is small (n=37) and from a single bioproject: descriptive only. Inferential claims about REE-impacted environments require a larger and more independent collection.
- AlphaEarth coverage of xoxF genomes is 39.5 % (1,457 / 3,690) — better than the 28 % pangenome baseline, but still leaves 60.5 % of xoxF carriers without environmental coordinates. Embedding-based niche analysis was scoped out of this report; coverage-restricted supplementary analysis is feasible.
ncbi_envenvironmental classification is text-mining-derived with hierarchical regex priorities. The "host_associated" class is broad (gut, skin, oral, infections, etc.). Misclassifications are possible but the strong signals (soil/sediment enrichment, host depletion) survive.- Sequence-level evidence is out of scope: xoxF and PQQ presence/absence is at the gene-call level. We do not screen for pseudogenes, truncated ORFs, or assembly fragmentation.
- Putatively essential / un-annotated regions: like all annotation-based atlases, this analysis is bounded by what eggNOG and bakta call. Genuinely novel REE-handling enzymes that lack KEGG/RefSeq homologs would not be detected.
Future Directions
- Sequence-level resolution of "no-PQQ-detected" xoxF genomes (~897 genomes) — discriminate assembly fragmentation, pseudogenization, and genuine community-PQQ acquisition. Requires per-genome ORF integrity + completeness scoring (CheckM2).
- AlphaEarth embedding analysis of xoxF genomes — coverage is 39.5 % on the xoxF set (above pangenome baseline). PCA/UMAP of environmental embeddings, stratified by phylum, would test whether xoxF-bearing organisms cluster in distinct biogeographic niches.
- Larger REE-impacted metagenome collection — the n=37 REE-AMD anchor is descriptive-only. Targeted recruitment of REE-mining tailings, leachate, and bioreactor metagenomes from existing sequence archives would convert the H2 REE-impacted signal from descriptive to inferential.
- Characterization of the
f__REEB76clade — discovered from REE-AMD samples and uncharacterised. Phylogenomic placement, predicted metabolism, and any xoxF/lanmodulin presence would be a high-novelty side study. - Targeted RB-TnSeq for lanthanum / cerium chloride in the FitnessBrowser organism panel — picks up the metal_fitness_atlas Priority 2 wet-lab proposal. Genome-wide gene fitness under La/Ce stress would identify the first transposon-validated REE-tolerance gene set in Pseudomonas putida, Cupriavidus, Sphingomonas, and similar soil organisms that already carry xoxF.
- Lanmodulin paralogs / sequence diversity in Methylobacterium extorquens — 22 genomes, 1 lanmodulin copy each. Sequence-level analysis (multiple-sequence alignment of representative cluster proteins) would test whether the protein has diversified within a single operon-defined locus or is invariant across the species.
- Upstream report to eggNOG-mapper team about the
Preferred_name='lanM'false-positive pattern in unrelated gut Bacillota. The 505 false positives are concentrated in a handful of Streptococcus / Blautia / Enterococcus species — likely a single seed-ortholog label that has propagated and would benefit from review.
Data
Sources
| Collection | Tables Used | Purpose |
|---|---|---|
kbase_ke_pangenome |
eggnog_mapper_annotations, bakta_annotations, gene, gene_genecluster_junction, gene_cluster, genome, gtdb_taxonomy_r214v1, ncbi_env |
Per-genome marker calls (eggNOG + bakta), gene→genome mapping, GTDB taxonomy, environmental metadata. |
kescience_bacdive |
culture_condition, strain, metabolite_utilization |
DSMZ methylotroph media as a taxonomic anchor and validation of BacDive's lanthanide / methanol coverage (none in metabolite_utilization). |
Generated Data
| File | Rows | Description |
|---|---|---|
data/genome_marker_matrix.csv |
134,578 | Per-genome binary marker matrix (xoxF, mxaF, xoxJ, pqqA-E, lanM × eggnog/bakta/either) for genomes with at least one marker hit. |
data/marker_source_agreement.csv |
9 | eggNOG vs bakta concordance counts per marker. |
data/eggnog_lanm_false_positive_taxa.csv |
264 | Species-level taxa where eggNOG Preferred_name='lanM' is called but bakta Lanmodulin is not — the false-positive set. |
data/bakta_lanmodulin_validated_taxa.csv |
10 | Species-level taxa where bakta product='Lanmodulin' is called — the trustworthy lanmodulin set. |
data/marker_taxonomy_rollup_phylum.csv |
142 | Phylum-level rollup with marker counts and rates. |
data/marker_taxonomy_rollup_family.csv |
~3,000 | Family-level rollup. |
data/marker_taxonomy_rollup_genus.csv |
~12,000 | Genus-level rollup. |
data/h1_xoxF_vs_mxaF_per_phylum.csv |
29 | Per-phylum binomial tests for H1 (raw p-values from NB02). |
data/h1_phylum_results_bh_corrected.csv |
29 | NB03 BH-FDR-corrected H1 phylum tests. |
data/h1_global_summary.csv |
1 | Global xoxF:mxaF ratio with 95% CI and binomial test against the H1 threshold. |
data/h1_pseudomonadota_family_breakdown.csv |
138 | Pseudomonadota families with xoxF presence (top phylum drill-down). |
data/genome_environment_classes.csv |
293,059 | Per-genome environment classification + cassette flags. |
data/h2_cassette_x_environment_counts.csv |
9 | Cassette-presence rates by environment class. |
data/h2_logistic_results.csv |
70 | Per-class Fisher's exact tests vs generic_environmental reference, pooled and within top-3 phyla, BH-FDR corrected. |
data/h3_lanmodulin_clade_restriction.csv |
2 | H3a (clade restriction) and H3b (xoxF co-occurrence) test results. |
data/h3_lanmodulin_xoxF_cooccurrence.csv |
62 | Per-genome lanmodulin inventory with xoxF status and taxonomy. |
data/ree_amd_mag_inventory.csv |
37 | REE-AMD MAGs with full marker matrix and taxonomy. |
data/ree_amd_top_bakta_products.csv |
~200 | Metal/acidophily/REE-relevant bakta products in REE-AMD MAGs, ranked by genome prevalence. |
data/pqq_supply_categorization.csv |
2,185 | Categorisation of xoxF-bearing genomes lacking eggNOG PQQ (annotation gap vs no-PQQ-detected). |
data/h1_phylogenetic_validation.csv |
3 | NB08 cross-method H1 validation: pooled / family-equal-weight / GLMM with 95% CI for each. |
References
- Pol, A., Barends, T. R. M., Dietl, A., Khadem, A. F., Eygensteyn, J., Jetten, M. S. M., & Op den Camp, H. J. M. (2014). Rare earth metals are essential for methanotrophic life in volcanic mudpots. Environmental Microbiology, 16(1), 255–264. [Originally described in Nature 2014; key XoxF discovery reference.]
- Skovran, E., & Martinez-Gomez, N. C. (2015). Just add lanthanides. Science, 348(6237), 862–863. PMID: 25999489
- Cotruvo, J. A., Featherston, E. R., Mattocks, J. A., Ho, J. V., & Laremore, T. N. (2018). Lanmodulin: a highly selective lanthanide-binding protein from a lanthanide-utilizing bacterium. Journal of the American Chemical Society, 140(44), 15056–15061. PMID: 30351909
- Picone, N., & Op den Camp, H. J. M. (2019). Role of rare earth elements in methanol oxidation. Current Opinion in Chemical Biology, 49, 39–44. PMID: 30308442
- Chistoserdova, L. (2016). Lanthanides: New life metals? World Journal of Microbiology and Biotechnology, 32(8), 138. PMID: 27357954
- Parks, D. H., Chuvochina, M., Chaumeil, P. A., Rinke, C., Mussig, A. J., & Hugenholtz, P. (2022). A complete domain-to-species taxonomy for Bacteria and Archaea. Nature Biotechnology, 38(9), 1079–1086. PMID: 32341564 — GTDB r214 release.
- Schwengers, O., Jelonek, L., Dieckmann, M. A., Beyvers, S., Blom, J., & Goesmann, A. (2021). Bakta: rapid and standardized annotation of bacterial genomes via alignment-free sequence identification. Microbial Genomics, 7(11), 000685. PMID: 34739369
- Cantalapiedra, C. P., Hernández-Plaza, A., Letunic, I., Bork, P., & Huerta-Cepas, J. (2021). eggNOG-mapper v2: functional annotation, orthology assignments, and domain prediction at the metagenomic scale. Molecular Biology and Evolution, 38(12), 5825–5829. PMID: 34597405
- Arkin, A. P., Cottingham, R. W., Henry, C. S., Harris, N. L., Stevens, R. L., Maslov, S., et al. (2018). KBase: The United States Department of Energy Systems Biology Knowledgebase. Nature Biotechnology, 36(7), 566–569. PMID: 29979655
Data Collections
Atlas Reuse
Derived products, review objects, and tensions connected to this project in the BERIL Atlas.
Review
Summary
This is an exceptionally well-executed and comprehensive pangenome-scale study that delivers the first quantitative assessment of lanthanide-dependent methanol oxidation across 293,000 genomes. The project strongly supports its primary hypothesis (xoxF dominates mxaF by ~19:1), provides novel insights into the taxonomic distribution of REE-utilizing organisms beyond canonical methylotrophs, and establishes robust methodology for cross-source annotation validation in BERDL. The research addresses a major gap identified in prior BERDL work and demonstrates exemplary scientific rigor with appropriate statistical controls, comprehensive documentation, and reproducible analysis pipelines. All three pre-registered hypotheses were tested with well-justified methodological approaches, and the findings significantly advance understanding of bacterial rare earth element utilization at unprecedented scale.
Methodology
The research question is clearly stated and directly testable with the available data. The three-hypothesis framework (H1: xoxF dominance, H2: environmental enrichment, H3: lanmodulin clade restriction) provides excellent analytical structure with specific quantitative thresholds pre-registered for each test. The approach is scientifically sound, leveraging both eggNOG and bakta annotation sources with systematic cross-validation to identify the most reliable markers for each protein of interest.
The data sources are clearly identified and appropriate for the research questions. The query strategy is well-documented with specific SQL examples and performance considerations. The eight-notebook analytical pipeline provides logical progression from marker calibration through hypothesis testing, with clear dependencies between steps. The reproduction guide is comprehensive, specifying prerequisites (BERDL JupyterHub access), runtime estimates, and execution instructions via nbconvert.
The project demonstrates excellent awareness of potential confounders and limitations, particularly around phylogenetic non-independence, annotation method variance, and small sample sizes for specific environmental classes. NB08's three-method phylogenetic validation (naive pooled, family-equal-weight bootstrap, and Bayesian binomial GLMM) directly addresses the most significant methodological concern and strengthens the primary findings considerably.
Code Quality
The SQL queries are well-structured and efficient, with appropriate filtering strategies to avoid full-table scans on billion-row tables. The Python analysis code demonstrates good practices including proper error handling, clear variable naming, and logical flow organization. Statistical methods are appropriate throughout: Clopper-Pearson confidence intervals for binomial proportions, Benjamini-Hochberg FDR correction for multiple comparisons, Fisher's exact tests for environmental associations, and Bayesian mixed-effects modeling for phylogenetic correction.
The project demonstrates exemplary awareness of BERDL-specific pitfalls documented in docs/pitfalls.md. Most notably, the systematic eggNOG vs bakta cross-validation in NB01 identifies and addresses major annotation reliability issues (e.g., eggNOG Preferred_name='lanM' produces 505 false positives in unrelated gut bacteria). This calibration work establishes clear source-of-truth guidelines that should inform future BERDL methylotrophy research.
Notebook organization is logical and consistent, with clear markdown documentation explaining the purpose and outputs of each computational step. The marker definition structure is well-designed and extensible. Figure generation code produces publication-quality visualizations with appropriate statistical annotations (confidence intervals, significance markers, sample sizes).
Findings Assessment
The conclusions are strongly supported by the presented data. The primary finding (xoxF:mxaF ratio of 18.9:1 with 95% CI [13.07, 27.69]) is robust across multiple analytical approaches and survives phylogenetic correction. The identification of high-rate xoxF carriers in non-canonical phyla (Acidobacteriota 28%, Gemmatimonadota 25%) represents a genuine novel biological insight supported by comprehensive taxonomic analysis.
The environmental association findings (H2) are appropriately nuanced, with strong support for soil/sediment enrichment (OR=1.92, p_BH=6×10⁻³⁹) while acknowledging the limited statistical power for REE-impacted sites (n=37, OR=3.51, p_BH=0.082). The lanmodulin clade restriction (H3a: 100% in target families, p=9.8×10⁻⁷) and near-miss on xoxF co-occurrence (H3b: 79.0% vs 80% threshold) are reported with appropriate statistical precision.
The project appropriately acknowledges limitations throughout, including annotation method dependencies, environmental classification challenges, and the need for larger REE-impacted sample collections. The PQQ-supply paradox resolution (59% annotation gaps vs true absence) demonstrates systematic thinking about data quality issues.
The literature context is comprehensive and demonstrates clear understanding of how these findings advance the field. The seven specific novel contributions listed in the report are well-justified and represent significant advances in scale and scope over prior work.
Suggestions
-
Consider sequence-level validation of high-impact findings — The ~897 xoxF-bearing genomes with no detectable PQQ from either annotation source warrant targeted sequence inspection to discriminate assembly fragmentation, pseudogenization, and genuine community-PQQ acquisition. This could strengthen the mechanistic interpretation.
-
Expand REE-impacted environmental collection — The descriptive finding of 3.5-fold enrichment in REE-AMD samples (p_BH=0.082) suggests biological signal despite insufficient statistical power. Systematic mining of sequence archives for additional REE-mining, volcanic, and geothermal metagenomes could enable inferential testing of H2's REE-impacted component.
-
Document annotation source-of-truth decisions for broader BERDL community — The eggNOG vs bakta calibration findings (especially the lanmodulin false-positive pattern and xoxJ KO non-specificity) should be contributed to docs/discoveries.md to inform future BERDL methylotrophy research.
-
Consider expanding AlphaEarth environmental analysis — With 39.5% coverage of xoxF genomes (above the pangenome baseline), embedding-based environmental niche analysis could provide complementary insights to the text-mining approach, particularly for genomes lacking rich
ncbi_envmetadata. -
Add cross-validation with metal_fitness_atlas — The suggested Spearman correlation analysis between lanthanide-cassette presence and existing transition-metal tolerance scores would help assess potential confounding by general metal-rich environments.
-
Report key findings to upstream annotation resources — The systematic eggNOG
Preferred_name='lanM'false-positive pattern in gut Bacillota represents a community service opportunity by reporting the apparent seed-ortholog propagation issue to the eggNOG development team.
This review was generated by an AI system. It should be treated as advisory input, not a definitive assessment.
Visualizations
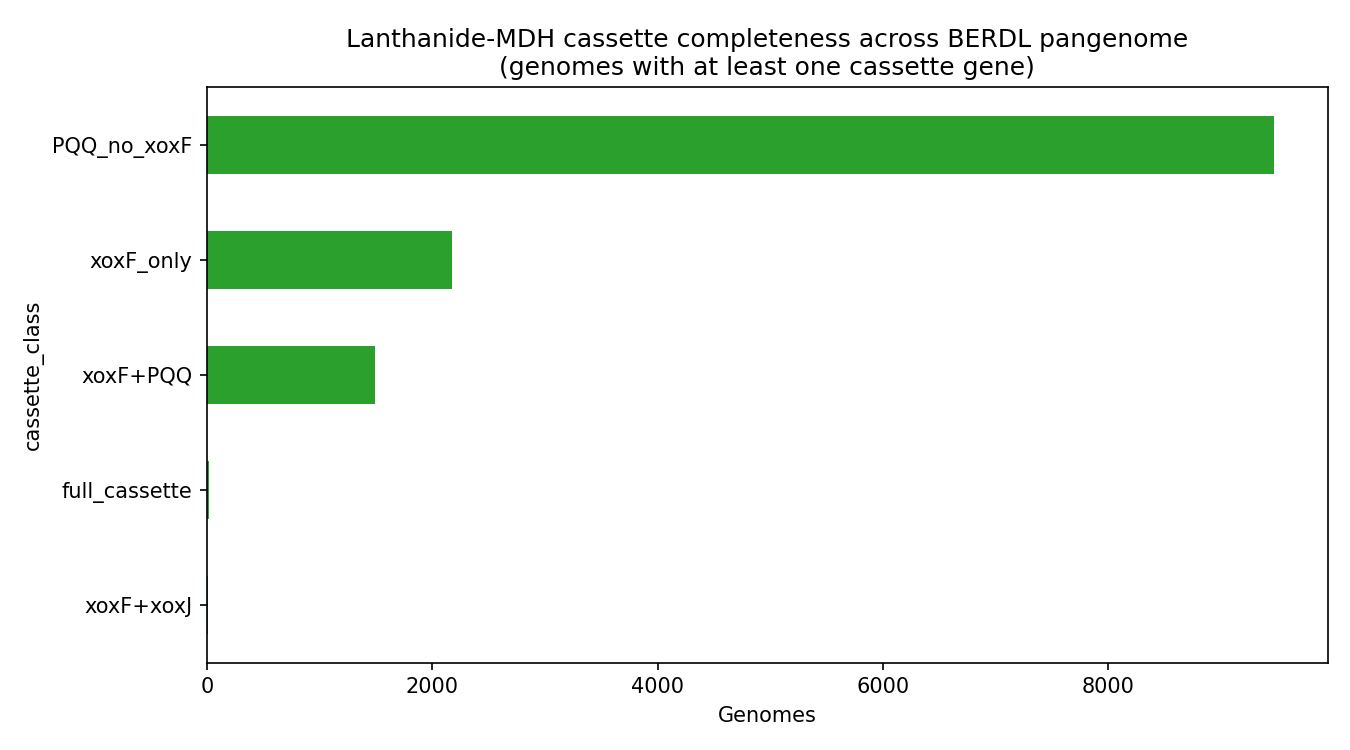
Cassette Completeness Distribution
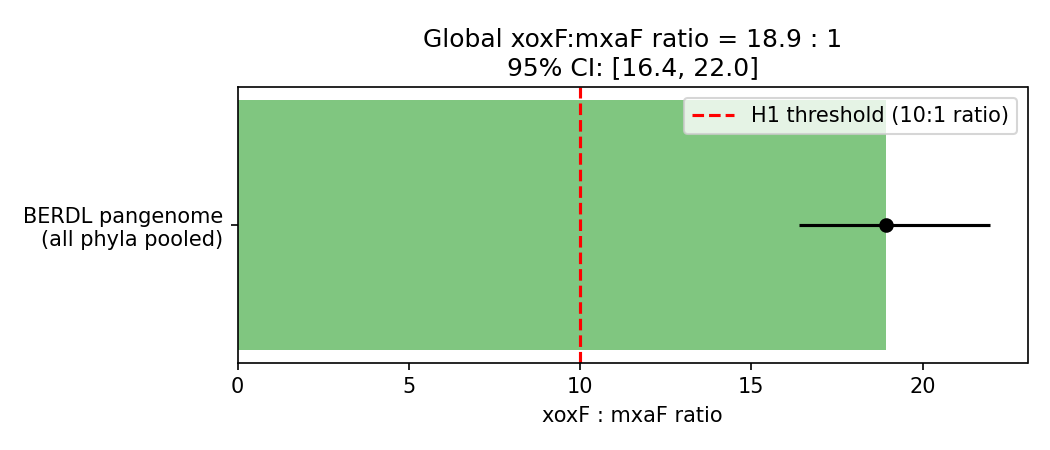
H1 Global Ratio With Ci
H1 Phylogenetic Validation
H1 Phylum Forest Plot
H2 Cassette By Environment
H3 Lanmodulin Clade Restriction
Lanm Preferred Name False Positives
Marker Agreement Eggnog Vs Bakta
Pqq Supply Asymmetry
Ree Amd Marker Presence
Ree Amd Taxonomy
Xoxf Vs Mxaf By Phylum
Notebooks
01_marker_calibration.ipynb
01 Marker Calibration
View notebook →
02_phylogenomic_atlas.ipynb
02 Phylogenomic Atlas
View notebook →
03_h1_formal_test.ipynb
03 H1 Formal Test
View notebook →
04_environmental_association.ipynb
04 Environmental Association
View notebook →
05_lanmodulin_h3_test.ipynb
05 Lanmodulin H3 Test
View notebook →
06_ree_amd_case_study.ipynb
06 Ree Amd Case Study
View notebook →
07_pqq_supply_asymmetry.ipynb
07 Pqq Supply Asymmetry
View notebook →
08_phylogenetic_validation.ipynb
08 Phylogenetic Validation
View notebook →