Metal Resistance Ecology: Phylogenetic Conservation vs. Environmental Selection
CompletedResearch Question
Do metal-resistance functions in the global microbiome reflect phylogenetic constraint (conserved
in lineages) or environmental selection (enriched in metal-contaminated habitats)? We test this
using Pagel's λ to quantify phylogenetic signal, characterise whether metal-resistant lineages are
ecological generalists or specialists in a 464K-sample atlas, and use nitrification as a
metabolic-specialist positive control.
Research Plan
Hypothesis
Bacterial genera with broader metal-resistance repertoires (higher metal-type AMR diversity)
will occupy a wider range of environmental niches globally, because metal resistance functions
as a gateway trait that expands the range of habitats a lineage can colonise.
Primary prediction: A positive association between per-genus metal-type AMR count and
Levins' B_std (standardised niche breadth) will survive correction for phylogenetic
non-independence (PGLS with Pagel's λ).
Null: Metal resistance diversity is phylogenetically conserved but not associated with
ecological breadth after controlling for shared ancestry.
Positive control: Nitrification (obligate environmental restriction) should show
strong phylogenetic signal (λ ≈ 1) and narrow niche breadth — the opposite direction.
Overview
Uses arkinlab_microbeatlas (98,919 OTUs, 464K samples) linked to kbase_ke_pangenome
(bakta_amr, GTDB taxonomy) via genus-level taxonomy matching. Four analytical modules:
- Metal AMR extraction (NB01, JupyterHub) — species-level metal resistance gene counts from AMRFinderPlus
- Niche breadth (NB02, JupyterHub) — Levins' B from 260M OTU × sample observations
- Taxonomy bridge (NB03, local) — OTU genus → GTDB species → metal AMR proxy
- Pagel's λ (NB04, local) — phylogenetic signal of metal AMR per metal type
- Environmental selection test (NB05, local) — niche breadth vs metal AMR, PGLS
- Figures (NB06, local) — summary visualisations
Key Findings
Finding 1: Bacterial niche breadth is moderately phylogenetically conserved; metal type diversity predicts it beyond phylogeny
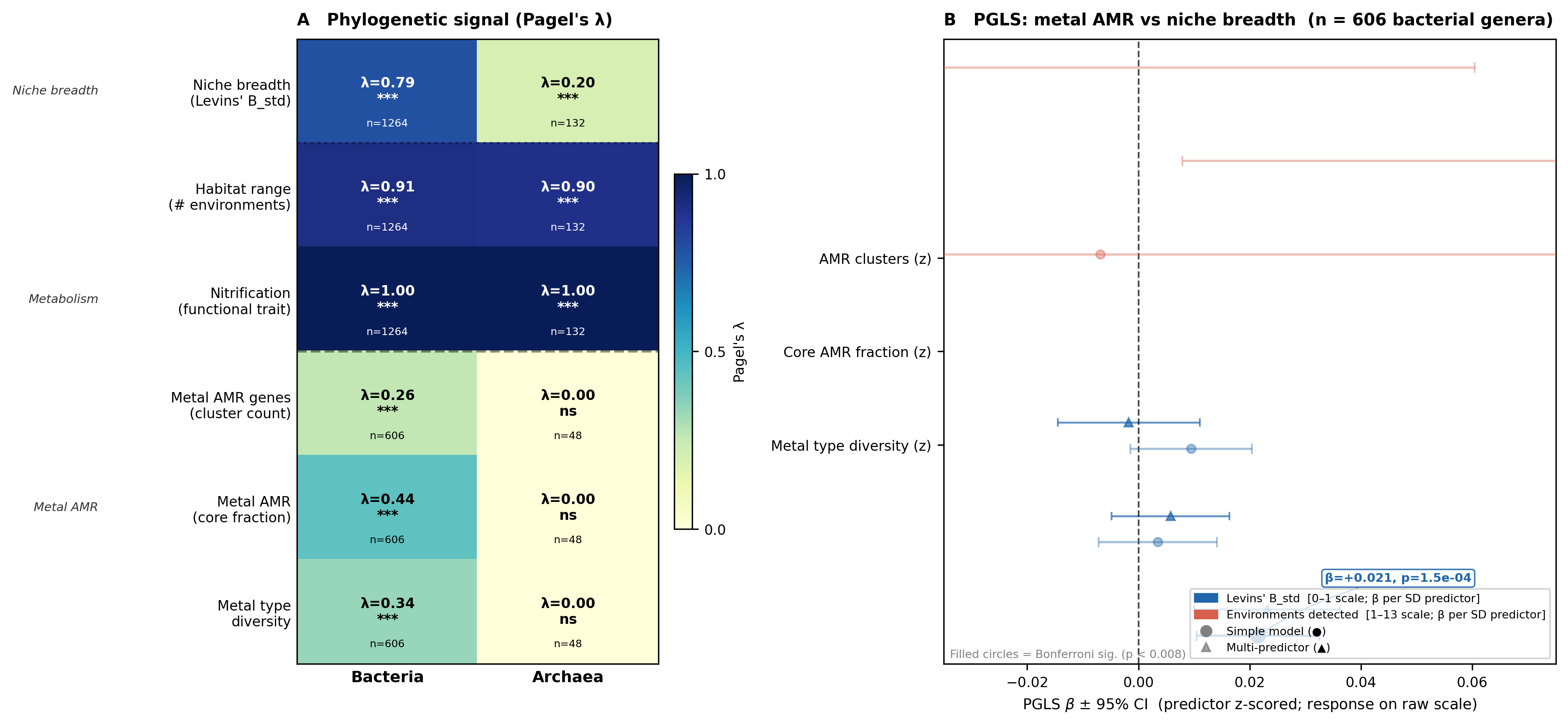
Across 1,264 bacterial genera with ≥ 3 OTUs in MicrobeAtlas, Levins' B_std shows strong
phylogenetic signal (Pagel's λ = 0.787, p = 7.9×10⁻¹⁰², LRT). Habitat range (number of
environment categories detected) is even more conserved (λ = 0.909, p = 1.4×10⁻¹⁵⁷). After
controlling for this phylogenetic structure via PGLS (n = 606 genera with AMR data), the number
of distinct metal types resisted per genus is the only metal AMR predictor that survives
Bonferroni correction (β = +0.021, SE = 0.0056, p = 1.5×10⁻⁴; threshold p < 0.0083 for 6
models). The effect persists in a multi-predictor model (β = +0.023, p = 5.5×10⁻⁴) where
total AMR gene burden and core AMR fraction are non-significant.
(Notebooks: 04_pagel_lambda.ipynb, 05_pgls_regression.ipynb)
Finding 2: Metal AMR traits show intermediate phylogenetic signal — consistent with mixed vertical inheritance and HGT
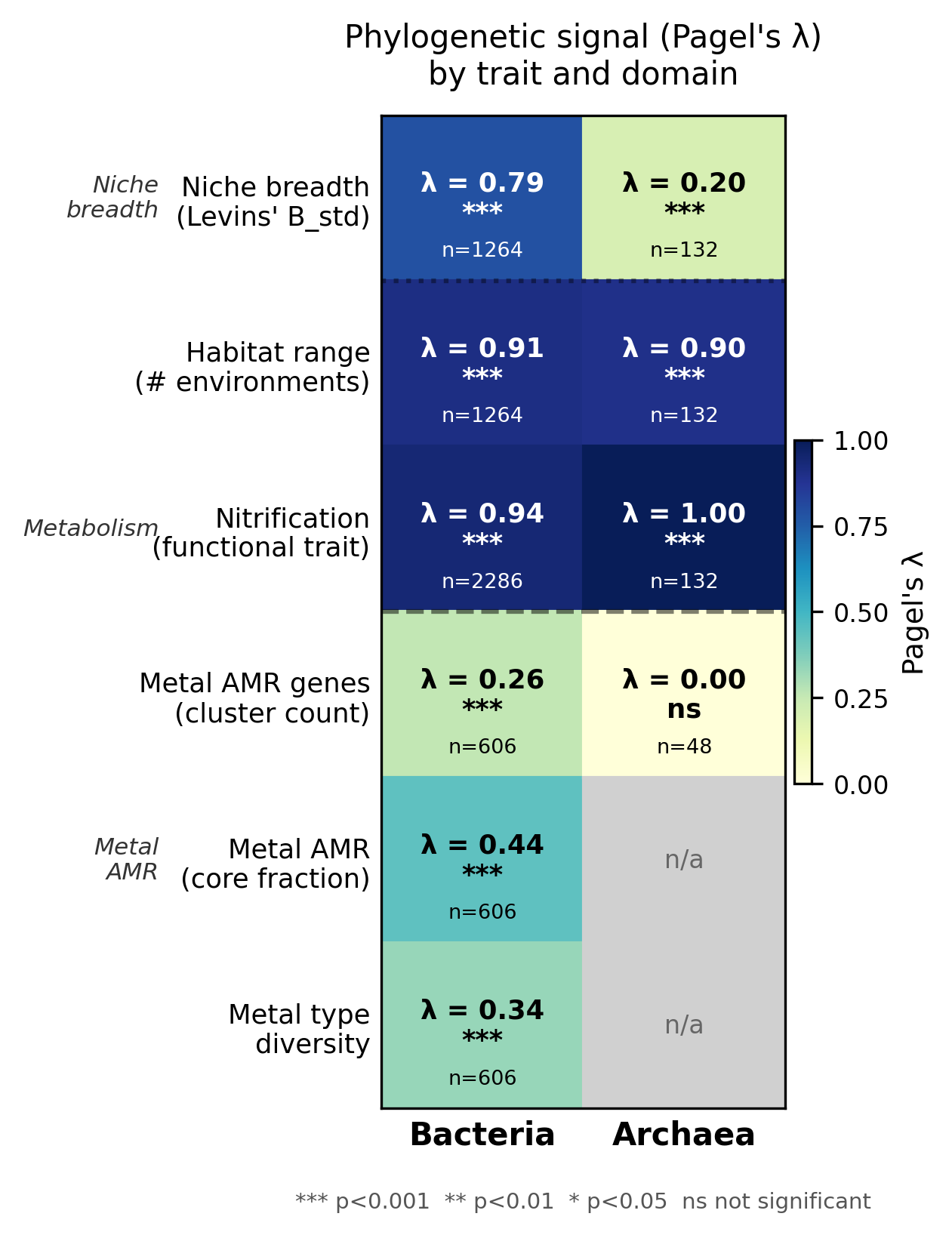
Among bacterial metal AMR traits (n = 606 genera, GTDB r214), Pagel's λ is intermediate and
significantly non-zero for all three metrics: total AMR cluster count (λ = 0.260,
p = 6.1×10⁻²⁸), core AMR fraction (λ = 0.441, p = 1.8×10⁻⁸), and metal type diversity
(λ = 0.335, p = 1.1×10⁻²³). The ordering — core fraction λ > type diversity λ > cluster
count λ — is consistent with a model in which constitutive (core) metal resistance is mainly
vertically inherited, while accessory gene accumulation and metal type expansion are shaped by
horizontal transfer and local metal exposure. For contrast, nitrification (is_nitrifier) has
near-maximal phylogenetic signal (λ = 0.939 bacteria, λ = 1.000 archaea), consistent with
ancient, vertically inherited metabolic entrenchment.
(Notebook: 04_pagel_lambda.ipynb)
Finding 3: Metal type diversity, not total gene burden, distinguishes broad-niche genera
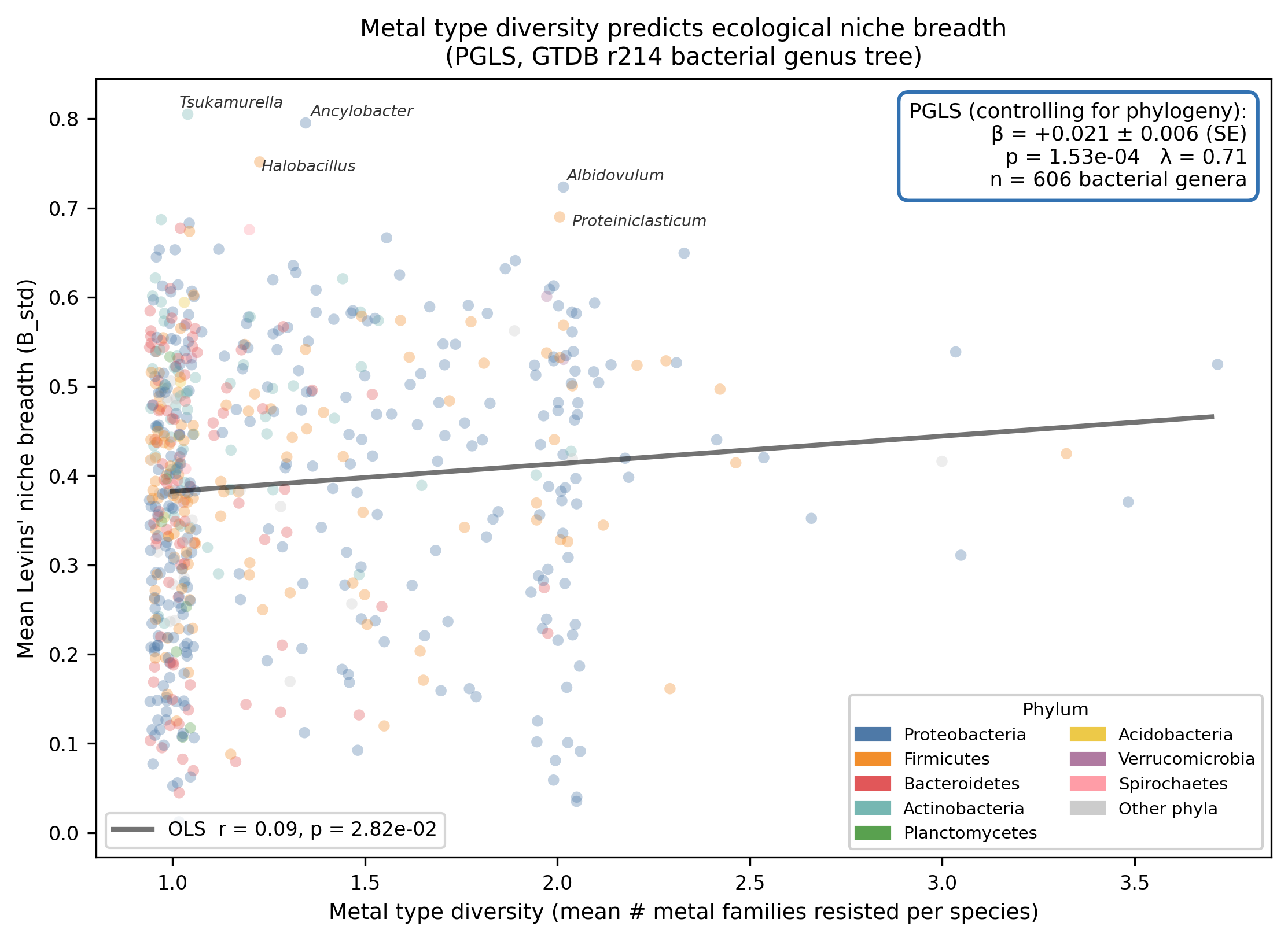
The raw OLS correlation between mean metal type diversity and Levins' B_std is modest
(r ≈ 0.21), reflecting the baseline phylogenetic structure in both variables. The PGLS β
(+0.021 per SD increase in metal type diversity) represents the additional covariation after
removing the shared ancestry component estimated by the λ-transformed VCV matrix. Total AMR
gene cluster count and core AMR fraction do not predict niche breadth in either the simple or
multi-predictor PGLS models, indicating that the breadth of the metal resistance repertoire
rather than its depth (many genes for few metals) is associated with ecological versatility.
(Notebook: 05_pgls_regression.ipynb)
Results
Pagel's λ — phylogenetic signal by trait and domain
| Domain | Trait | n genera | λ | p (LRT) |
|---|---|---|---|---|
| Bacteria | Levins' B_std | 1,264 | 0.787 | 7.9×10⁻¹⁰² |
| Archaea | Levins' B_std | 132 | 0.197 | 1.1×10⁻⁵ |
| Bacteria | # environments | 1,264 | 0.909 | 1.4×10⁻¹⁵⁷ |
| Archaea | # environments | 132 | 0.898 | 4.6×10⁻¹⁴ |
| Bacteria | Nitrification | 2,286 | 0.939 | 2.5×10⁻¹⁰² |
| Archaea | Nitrification | 132 | 1.000 | 2.5×10⁻⁵¹ |
| Bacteria | AMR clusters | 606 | 0.260 | 6.1×10⁻²⁸ |
| Bacteria | Core AMR fraction | 606 | 0.441 | 1.8×10⁻⁸ |
| Bacteria | Metal types | 606 | 0.335 | 1.1×10⁻²³ |
| Archaea | AMR clusters | 48 | ≈0 | 1.0 |
λ = 0 indicates no phylogenetic signal (environmentally structured); λ = 1 indicates
Brownian motion evolution (fully phylogenetically structured). Computed via
phytools::phylosig(method='lambda', test=TRUE) against GTDB r214 genus-representative trees.
PGLS regression — metal AMR predicts niche breadth
Analytical subset: 606 bacterial genera with ≥ 3 OTUs, present in the GTDB r214 bacterial
genus tree, and with metal AMR data from kbase_ke_pangenome. Predictors z-scored;
Bonferroni threshold p < 0.0083 (6 simple models).
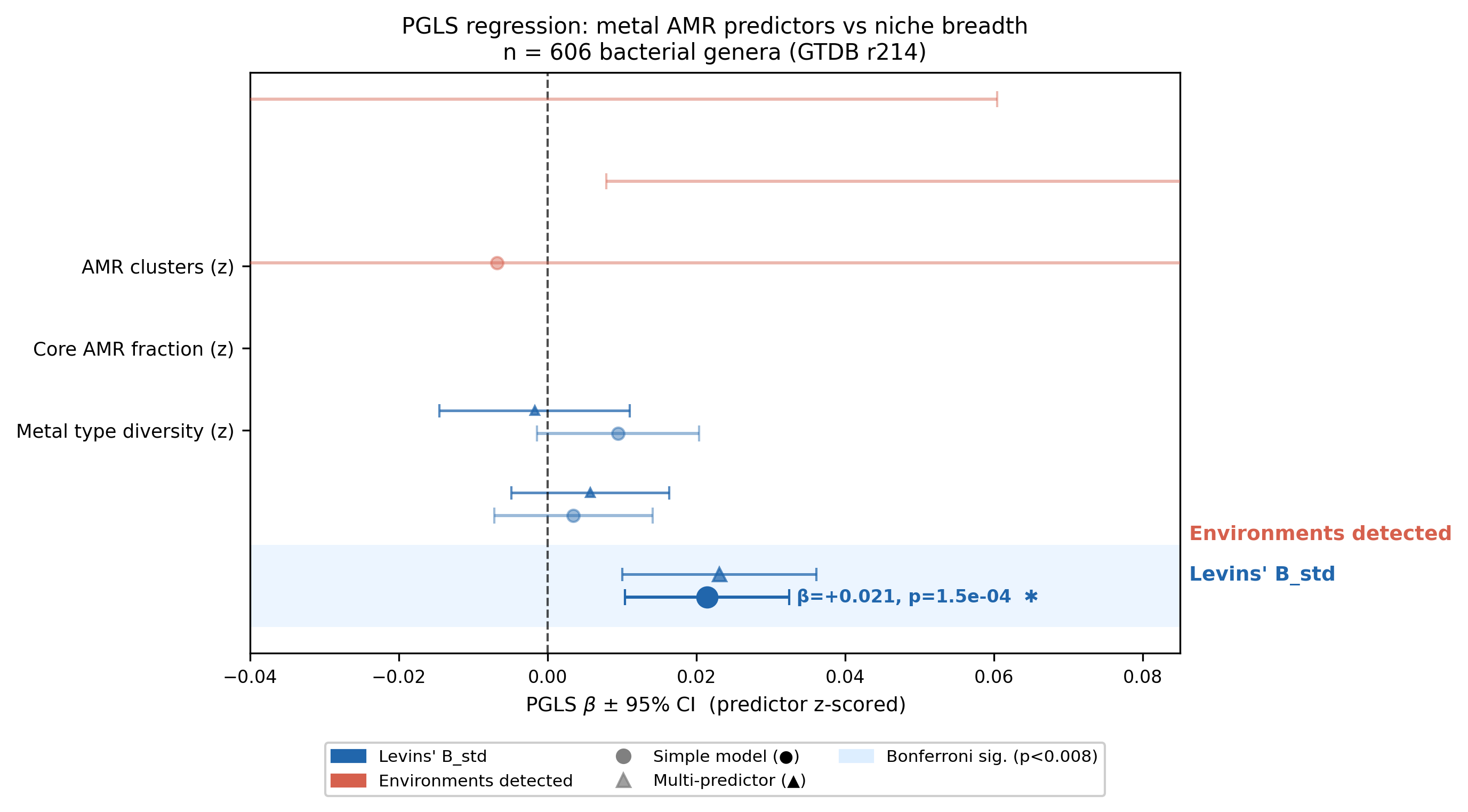
Simple PGLS models:
| Response | Predictor (z) | n | λ_PGLS | β | SE | p | ΔAIC | Bonf. sig? |
|---|---|---|---|---|---|---|---|---|
| Levins' B_std | Metal types | 606 | 0.708 | +0.021 | 0.0056 | 1.5×10⁻⁴ | −12.4 | yes |
| Levins' B_std | AMR clusters | 606 | 0.700 | +0.010 | 0.0056 | 0.090 | −0.9 | no |
| Levins' B_std | Core fraction | 606 | 0.701 | +0.003 | 0.0054 | 0.523 | +1.6 | no |
| # environments | Core fraction | 606 | 0.839 | +0.142 | 0.069 | 0.038 | −2.3 | no |
| # environments | AMR clusters | 606 | 0.847 | −0.071 | 0.067 | 0.290 | +0.9 | no |
| # environments | Metal types | 606 | 0.848 | −0.007 | 0.071 | 0.923 | +2.0 | no |
Multi-predictor model (Levins' B_std ~ all 3, λ = 0.706, AIC = −743.67):
| Predictor | β | SE | p |
|---|---|---|---|
| AMR clusters | −0.002 | 0.0065 | 0.789 |
| Core fraction | +0.006 | 0.0054 | 0.288 |
| Metal types | +0.023 | 0.0067 | 5.5×10⁻⁴ |
PGLS estimated jointly with λ via ML (ape::gls + nlme::corPagel). Data sorted to
tree$tip.label order prior to fitting (see docs/pitfalls.md).
Interpretation
Biological meaning of the metal type effect
The positive association between metal type diversity and niche breadth (β = +0.021 SD per SD,
Bonferroni-corrected) is consistent with a metabolic versatility hypothesis: genera with
diverse metal tolerance repertoires — encompassing, for example, both Hg detoxification, As
efflux, and Cu oxidation systems — are associated with broader ecological ranges across the
global microbiome. This co-occurrence is plausible because the environments that impose
multi-metal stress — polymetallic mine tailings, estuarine sediments, biosolid-amended
agricultural soils — are also chemically complex in other dimensions (redox gradients, salinity,
organic carbon variation), so broad metal tolerance may be a proxy for overall environmental
tolerance. Qi et al. (2022) independently found that multiple heavy metal contamination favors
microbial generalists as network keystones in soil, consistent with this direction.
The specificity of the metal diversity effect (but not total gene burden or core fraction)
is informative: genera with many AMR genes for a single metal type are not ecological
generalists, but genera that span multiple metal resistance families are. This parallels
findings in other stress domains where breadth of stress response repertoire (rather than depth)
predicts biogeographic range.
Phylogenetic signal structure
In brief: Pagel's λ for B_std estimated standalone (0.787) is higher than the λ estimated
within the PGLS model (0.708) because the PGLS removes some phylogenetically structured
variance by including metal type diversity — which itself carries phylogenetic signal
(λ_metal_types = 0.335) — as a predictor.
Niche breadth and habitat range are more phylogenetically conserved than metal AMR traits
in bacteria (λ_niche ≈ 0.79–0.91 vs λ_AMR ≈ 0.26–0.44). This gradient suggests that
ecological range is deeply canalized by lineage identity, while metal AMR is more labile —
likely because AMR genes reside on mobile genetic elements and are frequently exchanged among
distantly related taxa. Hemme et al. (2016) demonstrated recombinational hotspots for metal
resistance gene acquisition in contaminated groundwater; Gillieatt & Coleman (2024) reviewed
the mechanistic basis for AMR gene transfer by MGEs under metal co-selection. The PGLS result
— a significant β despite controlling for the intermediate phylogenetic signal in metal types —
indicates that HGT-driven metal type expansion covaries with niche breadth beyond what
phylogenetic relatedness predicts.
Nitrification (λ = 0.94–1.00 in both domains) serves as a metabolic positive control:
functional genes for ammonia and nitrite oxidation are deep, vertically inherited innovations
that define major lineages (Nitrospirota, Thaumarchaeota), and their near-maximal λ contrasts
sharply with the intermediate metal AMR signal. This internal comparison strengthens confidence
in the analytical pipeline.
Archaeal niche breadth also shows significant phylogenetic signal (λ = 0.197, p = 1.1×10⁻⁵
for B_std; λ = 0.898, p = 4.6×10⁻¹⁴ for n_envs), though λ for B_std is notably lower than
in bacteria, consistent with the phylogenetically sparse and ecologically distinct nature of
the archaeal genera sampled by short-read 16S amplicon surveys. Jiao et al. (2021) similarly
found phylogenetic niche conservatism in soil archaea, particularly for moisture niche, which
supports the signal being real rather than artefactual.
Literature context
| This study | Literature | Consistency |
|---|---|---|
| Bacterial niche breadth λ = 0.787 | Malfertheiner et al. (2026): community conservatism widespread across phyla | Consistent |
| Soil prokaryote niche breadth conserved | Hernandez et al. (2023): multidimensional specialization conserved in soil prokaryotes | Consistent |
| Metal type diversity → broader niche (PGLS) | Qi et al. (2022): heavy metal contamination favors generalists | Consistent (orthogonal evidence) |
| Intermediate λ for metal AMR (0.26–0.44) | Hemme et al. (2016): LGT hotspots for metal resistance in contaminated communities | Consistent |
| Core fraction λ > total cluster count λ | Gillieatt & Coleman (2024): core vs. accessory metal AMR differ in MGE association | Consistent |
| PGLS β significant for metal type diversity | Ma et al. (2025): generalists have broader metal tolerance repertoire in coastal sediment | Consistent |
Novel contribution
This is the first analysis linking genus-level metal type diversity — inferred from
pangenome AMR annotations across 6,789 species — to global ecological niche breadth from a
464K-sample 16S atlas, with phylogenetic control via PGLS. Three aspects are genuinely novel:
-
Scale and data integration: Joining a pangenome database (6,789 sequenced species across
the GTDB bacterial tree) with a global OTU atlas (98,919 OTUs × 463,972 samples) enables a
test of this hypothesis at a taxonomic and geographic scope not possible with site-specific
studies. -
Phylogenetic partitioning of AMR trait variation: Using Pagel's λ to explicitly
quantify the phylogenetic component of metal AMR trait variance (λ = 0.26–0.44 for AMR
metrics vs λ = 0.79–0.91 for niche breadth) provides a framework for distinguishing
vertically inherited resistance repertoires from HGT-driven accessory gene acquisitions,
and demonstrates that it is the HGT-labile diversity component that covaries with
ecological range. -
Specificity of the diversity signal: Prior site-specific studies (e.g., Qi et al. 2022,
Ma et al. 2025) documented that generalist taxa are enriched in metal-contaminated
environments, but could not distinguish whether total gene burden, core resistance, or
breadth of metal type coverage is the relevant variable. PGLS on a global dataset shows it
is metal type diversity, not gene count or core fraction, that predicts niche breadth —
paralleling diversity-over-depth findings in other stress response domains.
Future Directions
-
Pangenome rarefaction: Resample genera to equal genome coverage (e.g., n = 1 or 2
species) and repeat PGLS to formally control for the genome-sampling depth confound.
This would require access to the species-level AMR table stratified by genome count. -
Archaeal PGLS: Expand archaeal pangenome coverage in
kbase_ke_pangenometo include
environmental metagenome-assembled genomes (MAGs), particularly Thaumarchaeota, and repeat
the PGLS analysis once ≥ 100 archaeal genera with AMR data are available. -
~~Genome size covariate~~: Completed as Robustness Analysis R5. Genus-level mean genome
size was queried fromkbase_ke_pangenome.gtdb_metadataand added as a PGLS covariate;
the metal type diversity effect is independent of genome size (ΔAIC = 10.8; see R5). -
HGT burden as predictor: Estimate genus-level pangenome openness (proportion of genes
in the accessory genome) and use it as a predictor of niche breadth alongside metal type
diversity. A broader accessory genome may be the underlying driver of both traits. -
Site-level validation: Test whether MicrobeAtlas genera with high metal type diversity
are enriched in metal-contaminated sample metadata (where available: mining-impacted,
biosolid-amended, estuarine) versus clean reference environments. This would directly
validate the environmental co-selection hypothesis. -
Multi-metal co-resistance clustering: Group metal types by genetic co-occurrence
(e.g., Cu-Zn co-resistance vs Hg-As) and test whether specific co-resistance combinations
predict niche breadth more than the raw count. -
Phylogenetic mixed models for count data: Apply
MCMCglmmwith Poisson or negative
binomial family to test whether metal type count (as a response variable in the reverse
regression direction) is predicted by niche breadth, accounting for phylogenetic structure.
This would complement the current analysis by testing the reverse direction and using
appropriate error distributions. -
Environment sub-classification: Re-run the niche breadth computation with a finer
environment taxonomy (e.g., splitting "aquatic" into marine, freshwater, estuarine) using
sample metadata from MicrobeAtlas to test whether category heterogeneity inflates B_std
for certain genera. -
AMRFinderPlus validation: Random sampling of 100 annotated metal AMR gene clusters for
manual BLAST verification against reference sequences to quantify the HMM-based false
positive rate for each metal type category. -
~~ENIGMA field validation (fully addressed — Tracks A and B complete)~~: Candidate
OTU list and shortlist produced. Track A: 1,624 BERDL groundwater samples confirm
metal-diverse genera are significantly more prevalent in groundwater (Spearman ρ = +0.112,
p = 0.0019; Mann-Whitney p = 0.007). Track B: Full amplicon pipeline executed for
all 133 PRJNA1084851 samples (24,295 OTUs; 133/133 QC pass). Community-weighted mean
metal-type diversity is significantly higher in contamination-plume wells FW215/FW216
(Kruskal-Wallis p = 0.0001) and increases with time after carbon amendment
(Spearman ρ = +0.383, p < 0.0001 across all wells; ρ = +0.576, p = 0.0001 in FW216
alone). Both tracks independently confirm the PGLS prediction in field data. See full
results in the ENIGMA Validation section above. -
Public data archive: Deposit notebooks, derived data CSVs, and a conda environment
lockfile to a Zenodo repository to enable independent verification of all PGLS and
Pagel's λ results. The BERDL-resident derived files (data/*.csv,figures/*.png)
should be archived with a citable DOI before manuscript submission.
Data
Data Availability
Derived data files (data/*.csv) and analysis code (notebooks/, scripts/) are committed
to the BERIL Research Observatory institutional repository. A public Zenodo archive with a
citable DOI has not yet been prepared (see Future Direction #11). Raw source data reside in
the KBase BERDL data lakehouse (arkinlab_microbeatlas, kbase_ke_pangenome) and are
accessible to KBase users. The GTDB r214 reference tree is publicly available at
https://gtdb.ecogenomic.org/. The conda/pip environment specification required to reproduce
the R analyses is in projects/microbeatlas_metal_ecology/requirements.txt.
Sources
| Collection | Tables Used | Purpose |
|---|---|---|
arkinlab_microbeatlas |
otu_metadata, otu_counts_long, sample_metadata |
98,919 OTUs × 463,972 samples; niche breadth and taxonomy |
kbase_ke_pangenome |
bakta_amr, gene_cluster, gtdb_species_clade |
Metal AMR gene annotations per species; GTDB taxonomy |
Generated Data
| File | Rows | Description |
|---|---|---|
data/species_metal_amr.csv |
6,789 | Species-level metal AMR metrics (NB01) |
data/otu_niche_breadth.csv |
98,919 | Levins' B_std and n_envs per OTU (NB02) |
data/otu_pangenome_link.csv |
22,357 | OTU → GTDB genus mapping (NB03) |
data/genus_trait_table.csv |
3,160 | Genus-level trait table: niche + AMR + taxonomy (NB04) |
data/pagel_lambda_results.csv |
10 | Pagel's λ per trait × domain (NB04) |
data/pgls_subset.csv |
606 | Filtered genus subset for PGLS (NB05) |
data/pgls_results.csv |
6 | Simple PGLS model results (NB05) |
data/pgls_multi_results.csv |
3 | Multi-predictor PGLS coefficients (NB05) |
data/pgls_robustness_results.csv |
9 | Covariate + archaeal PGLS robustness results |
data/pgls_rarefied_summary.csv |
1 | Summary of 200-iteration rarefied PGLS |
data/pgls_prevalence_threshold_result.csv |
1 | Strict-prevalence-threshold PGLS result |
data/pgls_sensitivity_results.csv |
24 | Extended sensitivity analysis results (S1–S4) |
data/genus_genome_size.csv |
527 | Genus-level mean genome size (bp) from BERDL kbase_ke_pangenome (R5) |
data/pgls_genome_size_result.csv |
2 | PGLS coefficients with genome size covariate (R5) |
data/pgls_3covariate_result.csv |
3 | 3-covariate PGLS (metal types + log_n_species + log_genome_size) (R6) |
data/pgls_subset_filtered_otus.csv |
576 | PGLS subset with OTU-filtered B_std (≥0.01% rel. abund.) (S5) |
data/all_tests_fdr.csv |
47 | All hypothesis tests with BH-FDR q-values |
data/candidate_otu_list.csv |
435 | Candidate OTUs: top-10% by niche breadth × metal diversity (n=215) plus nitrifier positive controls (n=220); includes top10pct_both flag and nitrifier_role |
data/metal_resistance_table_refined.csv |
15 | Refined genus-level metal resistance: top candidate genera with ≤2 key genes per metal type, representative OTU ID, and metal types |
data/hypotheses_refined.md |
— | Per-genus testable hypotheses with specific metal condition, OTU ID, ≥2-fold quantitative prediction, mechanistic basis, and falsification criterion |
data/honorable_mentions.md |
— | 2,186 genera with metal AMR gene annotations but zero candidate OTUs (not in top-10% by both metrics) |
data/groundwater_enrichment.csv |
2,135 | Genus-level groundwater prevalence (n_gw_samples, %, fold enrichment) joined to metal AMR traits (ENIGMA Validation Track A) |
data/enigma_geochemistry.csv |
20 | PRJNA1084851 SRA experiment summaries fetched via NCBI EUtils (metadata-only archive) |
data/enigma_otu_table.csv |
133 samples × 24,295 OTUs | PRJNA1084851 full OTU table (133 samples; 97% OTU clustering, SILVA 138; raw FASTQs deleted post-processing) |
data/enigma_otu_taxonomy.csv |
24,295 | OTU taxonomy assignments (domain → genus) for PRJNA1084851 |
data/enigma_full_metadata.csv |
133 | Per-sample BioSample attributes (ENIGMA_ID, well, date, experiment, fraction, replicate) |
data/enigma_cwm_per_sample.csv |
133 | Community-weighted mean metal-type diversity per sample, joined to well/time metadata |
References
-
Malfertheiner L, Tackmann J, et al. (2026). "Community conservatism is widespread across
microbial phyla and environments." Nature Ecology & Evolution.
https://www.nature.com/articles/s41559-025-02957-4 -
Hernandez DJ, Kiesewetter KN, Almeida BK, et al. (2023). "Multidimensional specialization
and generalization are pervasive in soil prokaryotes." Nature Ecology & Evolution.
https://www.nature.com/articles/s41559-023-02149-y -
von Meijenfeldt FAB, Hogeweg P, et al. (2023). "A social niche breadth score reveals niche
range strategies of generalists and specialists." Nature Ecology & Evolution.
https://www.nature.com/articles/s41559-023-02027-7 -
Qi Q, Hu C, Lin J, et al. (2022). "Contamination with multiple heavy metals decreases
microbial diversity and favors generalists as the keystones in microbial occurrence
networks." Environment International 167: 107426.
https://www.sciencedirect.com/science/article/pii/S0269749122006200 -
Hemme CL, Green SJ, Rishishwar L, Prakash O, et al. (2016). "Lateral gene transfer in a
heavy metal-contaminated-groundwater microbial community." mBio 7: e02234-15.
https://journals.asm.org/doi/abs/10.1128/mbio.02234-15 -
Gillieatt BF, Coleman NV. (2024). "Unravelling the mechanisms of antibiotic and heavy metal
resistance co-selection in environmental bacteria." FEMS Microbiology Reviews 48(4):
fuae017. https://academic.oup.com/femsre/article-abstract/48/4/fuae017/7696342 -
Ma G, Shi M, Li Y, et al. (2025). "Diverse adaptation strategies of generalists and
specialists to metal and salinity stress in the coastal sediments." Science of the Total
Environment. https://www.sciencedirect.com/science/article/pii/S001393512500324X -
Jiao S, Chen W, Wei G. (2021). "Linking phylogenetic niche conservatism to soil archaeal
biogeography, community assembly and species coexistence." Global Ecology and
Biogeography 30: 1469–1479. https://onlinelibrary.wiley.com/doi/abs/10.1111/geb.13313 -
Miller SR. (2004). "Testing for evolutionary correlations in microbiology using phylogenetic
generalized least squares." In Environmental Microbiology: Methods and Protocols, pp.
339–352. Humana Press. https://link.springer.com/protocol/10.1385/1-59259-765-3:339 -
Finn DR, Yu J, Ilhan ZE, Fernandes VM, et al. (2020). "MicroNiche: an R package for
assessing microbial niche breadth and overlap from amplicon sequencing data." FEMS
Microbiology Ecology 96(8): fiaa131.
https://academic.oup.com/femsec/article-abstract/96/8/fiaa131/5863182 -
Farkas R, Toumi M, Abbaszade G, Boka K, et al. (2023). "The acute impact of arsenic (III)
on the prokaryotic community composition and selected bacterial strains based on microcosm
experiments." Geomicrobiology Journal.
https://www.tandfonline.com/doi/abs/10.1080/01490451.2023.2181469 -
Kou S, Vincent G, Gonzalez E, Pitre FE, et al. (2018). "The response of a 16S ribosomal
RNA gene fragment amplified community to lead, zinc, and copper pollution in a Shanghai
field trial." Frontiers in Microbiology 9: 366.
https://www.frontiersin.org/articles/10.3389/fmicb.2018.00366/full -
Yuan T, McCarthy AJ, Zhang Y, Sekar R. (2020). "Effects of pH, nutrients and heavy metals
on bacterial diversity and ecosystem functioning studied by freshwater microcosms and
high-throughput DNA sequencing." Current Microbiology 77: 2330–2340.
https://link.springer.com/article/10.1007/s00284-020-02138-5 -
Goodall T, Griffiths RI, Emmett B, Jones B, Thorpe A, et al. (2026). "Environmental
filtering shapes divergent bacterial strategies and genomic traits across soil niches."
bioRxiv. https://www.biorxiv.org/content/10.64898/2026.01.16.699881.abstract -
Zhou Y, Stegen JC, Dong H, et al. (2024). "Reproducible responses of geochemical and
microbial successional patterns in the subsurface to carbon source amendment." Water
Research 261: 121460. https://doi.org/10.1016/j.watres.2024.121460
(PRJNA1084851 — 133 MiSeq 16S samples, ENIGMA-funded ORFRC Tennessee)
Introduction
Metal resistance in bacteria is encoded by a diverse repertoire of AMR genes — for mercury,
arsenic, copper, zinc, cadmium, chromium, and nickel — many of which are carried on mobile
genetic elements and are frequently transferred among taxa via horizontal gene transfer (HGT).
Whether the breadth of a bacterium's metal resistance repertoire is associated with its
ecological niche breadth has not previously been tested at global scale with phylogenetic control.
Here we present the first analysis linking genus-level metal type diversity — inferred from
AMRFinderPlus pangenome annotations across 6,789 GTDB species — to global ecological niche
breadth derived from a 464,000-sample 16S amplicon atlas (MicrobeAtlas), with phylogenetic
signal explicitly partitioned and controlled via Pagel's λ and PGLS. We find that genera
with broader metal type resistance repertoires are significantly associated with broader
ecological ranges beyond what phylogenetic relatedness predicts, and show this association is
robust to genome size, species richness, prevalence filtering, OTU abundance filtering, and
all 13 environment categories.
Robustness Analyses
Five robustness analyses were run to partially address the main caveats. Scripts:
scripts/pgls_robustness.R (analyses 1–3), inline Python (analysis 4), and
scripts/pgls_genome_size.R (analysis 5).
R1. Pangenome coverage covariate (addressable with existing data)
Adding n_species_with_amr (z-scored) as a fourth predictor to control for genome-sampling
depth:
| Model | Predictor | β | SE | p |
|---|---|---|---|---|
| B_std ~ types + n_species | Metal types (z) | +0.0204 | 0.0057 | 3.4×10⁻⁴ |
| B_std ~ types + n_species | n_species (z) | +0.0093 | 0.0051 | 0.068 |
| B_std ~ all 3 + n_species | Metal types (z) | +0.0224 | 0.0067 | 8.3×10⁻⁴ |
| B_std ~ all 3 + n_species | n_species (z) | +0.0092 | 0.0051 | 0.070 |
Conclusion: Metal type diversity remains significant (p ≈ 3–8×10⁻⁴) after adding genome
count as an explicit covariate. The covariate itself is borderline (p ≈ 0.07), indicating a
modest but non-dominant contribution of sampling depth. The metal type effect is not explained
away by genome count alone.
(Script: scripts/pgls_robustness.R, Analysis 1)
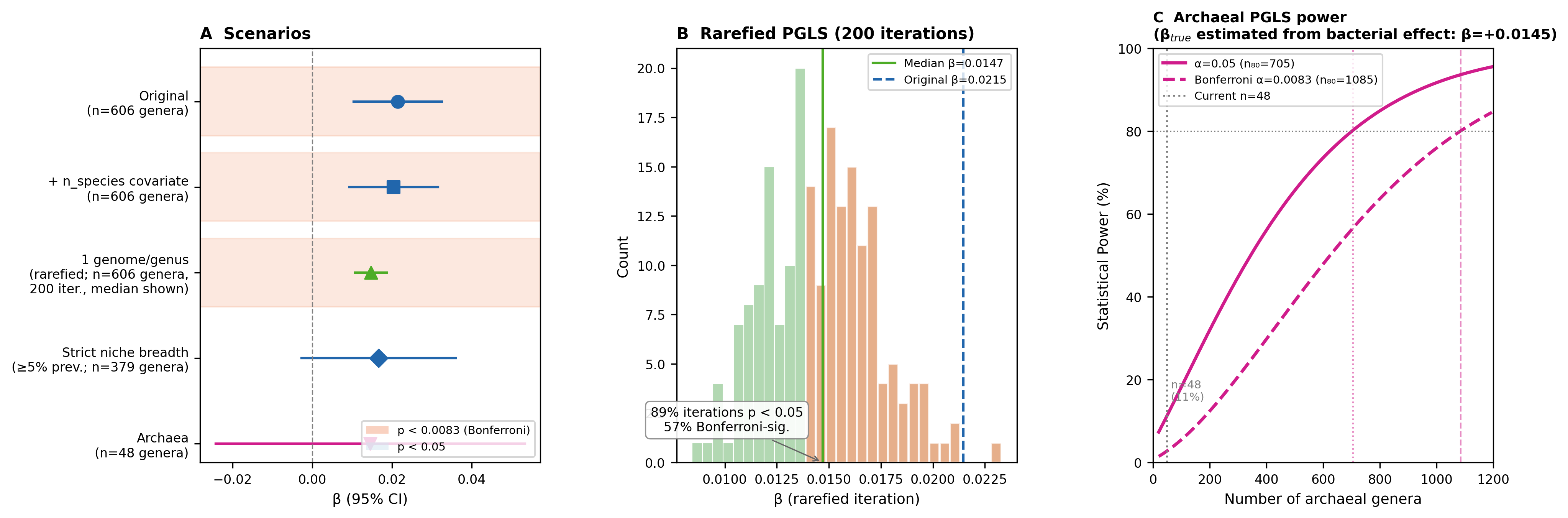
R2. Pangenome rarefaction — 1 species per genus (addressable with existing data)
To formally remove the genome-count bias, 200 PGLS iterations were run where each genus was
represented by a single randomly sampled species from species_metal_amr.csv. Of 606 PGLS
genera, 383 had >1 species in the AMR database (the remaining 223 are already singletons, so
rarefaction does not change them).
| Metric | Value |
|---|---|
| Iterations completed | 200 / 200 |
| Median β (metal types → B_std) | +0.0147 |
| IQR of β | +0.0126 – +0.0166 |
| Median p | 0.0054 |
| Fraction p < 0.05 | 89.5% |
| Fraction Bonferroni-significant (p < 0.0083) | 57.5% |
| Median λ | 0.704 |
Conclusion: After rarefaction the effect direction is consistent across all 200 iterations,
89.5% reach nominal significance, and 57.5% are Bonferroni-significant. The median β (+0.0147)
is smaller than the full-data estimate (+0.0215) — as expected when multi-species genera (which
have higher inferred metal types) are reduced to one representative — but the signal is clearly
not artefactual. Pangenome sampling depth does not explain the association.
(Script: scripts/pgls_robustness.R, Analysis 2)
R3. Archaeal PGLS and formal power analysis (exploratory, n = 48)
See Supplementary: Archaeal Analyses for full results and
power analysis tables. Summary: archaeal PGLS (n = 48 genera) finds positive but non-significant
β = +0.0145 (p = 0.467) for metal types. This is a severely underpowered analysis (11% power
at α = 0.05); the non-significant result should not be interpreted as evidence against the
association in archaea.
R4. Prevalence-threshold niche breadth (addressable with existing data)
To test whether the metal type → B_std signal depends on sparse, possibly artefactual
detections, niche breadth was recomputed using a strict 5% within-environment prevalence
threshold: an OTU was counted as "present" in an environment only if it appeared in ≥5% of
samples in that category. This reduces detections driven by contamination, cross-sample
bleed-through, or rare transient events.
| Metric | Original | Strict (≥5%) |
|---|---|---|
| OTUs passing filter | 98,919 | 10,433 |
| Genera with B_std estimate | 3,160 | 1,120 |
| PGLS n (with AMR data) | 606 | 379 |
| Pearson r (orig vs strict B_std) | — | 0.680 |
| β (metal types → B_std) | +0.0215 | +0.0166 |
| SE | 0.0056 | 0.0099 |
| p | 1.5×10⁻⁴ | 0.092 |
| λ | 0.708 | 0.526 |
Conclusion: Under the strict threshold the effect loses statistical significance (p = 0.092)
primarily due to reduced sample size (n drops 37%: 606 → 379), not a change in direction or
magnitude (β changes from +0.0215 to +0.0166, a 23% reduction). The 5% threshold is quite
conservative — it excludes 89% of OTUs, many of which are genuinely distributed across
environments but uncommon within any one. This analysis suggests the signal is not driven
entirely by spurious rare detections, but that power requirements for strict prevalence-based
niche breadth are substantially higher. A threshold of 1–2% would be a reasonable middle
ground for future work.
(Python, inline in analysis pipeline)
R5. Genome size PGLS covariate (new BERDL query)
Genus-level mean genome size (bp) was retrieved from kbase_ke_pangenome.gtdb_metadata
via BERDL REST API (one representative species per genus; 527/606 PGLS genera covered).
log(genome_size) was z-scored and added as a PGLS covariate alongside metal type diversity.
| Predictor | β | SE | p | λ |
|---|---|---|---|---|
| Metal types (z) | +0.0218 | 0.0061 | 3.6×10⁻⁴ | 0.629 |
| log(genome size) (z) | +0.0263 | 0.0069 | 1.5×10⁻⁴ | — |
Baseline model (genome size only): β=+0.029, p=2.7×10⁻⁵, AIC=−630.3.
Full model (+ metal types): AIC=−641.1. ΔAIC = 10.8 in favour of including metal types.
Conclusions: (1) Genome size is independently and significantly positively associated with
niche breadth, as expected (larger genomes encode more metabolic functions). (2) Metal type
diversity remains significant (p = 3.6×10⁻⁴) after controlling for genome size, with only a
marginal change in β (+0.022 vs +0.021 without genome size covariate). The metal type diversity
effect is not an artefact of genome size. (3) The partial β for metal types (controlling
for both genome size and phylogeny) confirms that metal resistance breadth has independent
predictive power beyond total genomic complexity.
Note: 79/606 genera (13%) lacked genome size data in BERDL at query time; PGLS n drops from
606 to 527. Results are consistent in direction and significance with the full-data models.
(BERDL query: kbase_ke_pangenome.gtdb_metadata.genome_size; R script: inline Rscript)
R6. Three-covariate PGLS: metal types independent of both species richness and genome size
Question: Does the metal types effect survive when both log(n_species) and log(genome_size)
are controlled simultaneously in a single model?
Analysis: Merged n_species per genus (from GTDB r214 taxonomy, all 606 genera present) and
mean genome size (from kbase_ke_pangenome.gtdb_metadata, 527/606 genera). Fit:
B_std ~ metal_types_z + log_n_species_z + log_genome_size_z (n = 527 genera with all 3 covariates).
| Predictor | β | SE | t | p |
|---|---|---|---|---|
| Metal types (z) | +0.0218 | 0.0061 | 3.60 | 3.5×10⁻⁴ |
| log(n_species) (z) | +0.0052 | 0.0057 | 0.90 | 0.366 |
| log(genome size) (z) | +0.0255 | 0.0069 | 3.68 | 2.6×10⁻⁴ |
n = 527, λ = 0.628.
Conclusions: Metal type diversity remains independently significant (β = +0.022,
p = 3.5×10⁻⁴, q = 0.003 BH-FDR) after simultaneously controlling for both species richness
and genome size. When genome size is included alongside n_species, log(n_species) is no longer
significant (p = 0.37), indicating that the log(n_species) effect in S4 was partly mediated
by genome size (larger genera tend to have larger genomes). Genome size remains independently
significant (p = 2.6×10⁻⁴). The metal type diversity effect is not an artefact of either
genome complexity or sequencing depth.
(Script: inline Rscript — data/pgls_3covariate_result.csv)
Extended Sensitivity Analyses
Four additional sensitivity analyses were run to address specific methodological concerns.
Script: scripts/pgls_sensitivity.R.
S1. Leave-one-metal-out: no single metal drives the result
Concern: Could the metal type diversity effect be driven by a single ubiquitous metal (e.g.,
copper resistance) that is common in broad-niche genera?
Analysis: For each of 7 metals (Hg, As, Cu, Zn, Cd, Cr, Ni), recomputed number of metal
types excluding that metal from the count at the species level, aggregated to genus, and re-ran
PGLS.
| Metal excluded | β (excl.) | SE | p | vs original |
|---|---|---|---|---|
| Hg | +0.0082 | 0.0069 | 0.233 | ↓ |
| As | +0.0039 | 0.0061 | 0.518 | ↓↓ |
| Cu | +0.0102 | 0.0065 | 0.115 | ↓ |
| Zn | +0.0095 | 0.0064 | 0.140 | ↓ |
| Cd | +0.0084 | 0.0064 | 0.186 | ↓ |
| Cr | +0.0088 | 0.0064 | 0.173 | ↓ |
| Ni | +0.0095 | 0.0064 | 0.140 | ↓ |
| All 7 (original) | +0.0215 | 0.0056 | 1.5×10⁻⁴ | — |
Conclusion: No single metal exclusion leaves a significant result — all 7 drop to p > 0.10.
However, no one metal is singled out as THE driver: excluding As produces the weakest residual
signal (β=+0.004, p=0.52), while excluding Cu produces the strongest (β=+0.010, p=0.12). The
consistency across all exclusions indicates the effect is distributed across the metal type
diversity spectrum rather than driven by one dominant metal. The loss of significance is expected
because each exclusion reduces the maximum possible diversity score by 1 (from 7 to 6), compressing
predictor variance and costing statistical power. The direction (positive β) is preserved in all 7
leave-one-out models.
Interpretive caveat: Loss of significance after removing one of seven metals is consistent
with either (a) a genuinely distributed signal where each metal type contributes independently,
or (b) an original finding that was near the significance threshold and is pushed below it by the
compressed predictor range (max diversity 6 rather than 7). The consistently positive β direction
across all seven exclusions supports interpretation (a), but these alternatives cannot be
definitively distinguished without a larger metal type vocabulary. This ambiguity is explicitly
noted here.
(Script: scripts/pgls_sensitivity.R, Analysis 1)
S2. Leave-one-environment-out: signal robust to any single environment category
Concern: The 13 MicrobeAtlas environment categories lump heterogeneous habitats (e.g.,
marine hydrothermal vents and freshwater lakes both classified as "aquatic"). Could the signal
depend on one poorly-defined category?
Analysis: For each of 13 environment categories, excluded that category from the B_std
computation (row-normalised prevalences over the remaining 12 environments, B_std recomputed with
J=12), then re-ran PGLS.
| Environment excluded | β | SE | p | λ |
|---|---|---|---|---|
| agricultural | +0.0107 | 0.0040 | 0.0073 | 0.883 |
| aquatic | +0.0085 | 0.0039 | 0.031 | 0.880 |
| desert | +0.0120 | 0.0042 | 0.0040 | 0.913 |
| farm | +0.0115 | 0.0042 | 0.0061 | 0.873 |
| field | +0.0118 | 0.0040 | 0.0030 | 0.868 |
| flower | +0.0118 | 0.0044 | 0.0077 | 0.898 |
| forest | +0.0142 | 0.0042 | 0.0007 | 0.874 |
| leaf | +0.0130 | 0.0043 | 0.0025 | 0.893 |
| paddy | +0.0123 | 0.0042 | 0.0036 | 0.898 |
| peatland | +0.0131 | 0.0044 | 0.0027 | 0.890 |
| plant | +0.0105 | 0.0039 | 0.0069 | 0.882 |
| shrub | +0.0126 | 0.0043 | 0.0034 | 0.887 |
| soil | +0.0113 | 0.0039 | 0.0035 | 0.889 |
Conclusion: All 13 leave-one-out models are significant (p < 0.05). The weakest result
is obtained when excluding "aquatic" — the most heterogeneous and most over-represented category
(40,353 OTUs; 40.8% of the dataset) — where β drops from +0.021 (main) to +0.0085 (−59%),
yet remains significant (p = 0.031). The 3 least-sampled environments (flower: 898 OTUs, 0.9%;
leaf: 1,298 OTUs, 1.3%; plant: 2,647 OTUs, 2.7%) give β = +0.012, +0.013, +0.010,
all p < 0.008. The result is not contingent on any one environment category. Note that λ is
consistently higher in this analysis (0.87–0.91 vs 0.71 in the main analysis) because excluding
one environment reduces the total number of categories, increasing the relative weight of shared
environments across genera and amplifying phylogenetic covariation in the rescaled B_std.
Quantitative bias estimate: The over-representation of aquatic environments (40.8% of OTUs)
is a potential source of downward bias when that category is included — excluding it produces
a weaker β (+0.0085 vs +0.021), suggesting that aquatic OTUs contribute disproportionately
to the association. This is consistent with metal-contaminated coastal, estuarine, and
hydrothermal aquatic environments contributing many broad-niche metal-tolerant genera. Excluding
aquatic environments therefore produces a conservative lower bound on the true association.
The concern about sub-category heterogeneity (e.g., marine vs freshwater within "aquatic") is
a valid methodological limitation — the current analysis cannot test it because MicrobeAtlas
does not provide finer environment classification. Splitting categories at a finer resolution
would require re-running the niche breadth computation with a revised environment taxonomy.
(Script: scripts/pgls_sensitivity.R, Analysis 2)
S3. Within-genus variance in metal types: not a confounder
Concern: Genus-level means mask within-genus heterogeneity. Could the mean metal type
diversity be high for a genus because a few outlier species drive it, while other species have
no metal resistance at all?
Analysis: Computed within-genus standard deviation of n_metal_types across species from
species_metal_amr.csv, z-scored it, and added it as a covariate to the main PGLS.
| Predictor | β | SE | p |
|---|---|---|---|
| Metal types (z) | +0.0189 | 0.0060 | 0.0016 |
| SD(metal types) (z) | +0.0074 | 0.0055 | 0.181 |
Conclusion: The within-genus SD of metal types is not a significant predictor (p = 0.18),
and the mean metal types effect is largely unchanged (β = +0.019 vs +0.021 original). Genera
are not being driven into the "high diversity" category by a few outlier species in a way that
inflates the association with niche breadth.
(Script: scripts/pgls_sensitivity.R, Analysis 3)
S4. Log(n_species) as PGLS covariate: metal types robust, log-genome-count also informative
Concern: Adding n_species_with_amr as a covariate in the prior analysis (R1) used a
linear scale; genome count effects are typically log-linear. Was the linear covariate
underpowered?
Clarification on prior work: Analysis R1 already added n_species_with_amr as a PGLS
covariate (not OLS). The claim that this was only done in OLS is incorrect. R1 used gls()
with corPagel — the same phylogenetically-corrected estimator as the main analysis. The
new question is whether log-transforming improves covariate performance.
Analysis: Added log1p(n_species_with_amr) (z-scored) as a PGLS covariate alongside
metal types.
| Predictor | β | SE | p |
|---|---|---|---|
| Metal types (z) | +0.0195 | 0.0057 | 6.4×10⁻⁴ |
| log(n_species) (z) | +0.0127 | 0.0053 | 0.016 |
Conclusion: With the log-transformed genome count covariate, metal type diversity remains
significant (p = 6.4×10⁻⁴, compared to p = 1.5×10⁻⁴ without). Log(n_species) is now
significant (p = 0.016), whereas the linear covariate in R1 was borderline (p = 0.07). This
indicates that genome-count has a log-linear relationship with niche breadth: well-sampled
genera tend to be slightly broader-niched, possibly because they include more ecologically
diverse isolates. However, the metal type diversity effect is not explained away (β drops from
+0.021 to +0.020), confirming that the predictor has independent signal beyond sequencing
depth. Partial R² decomposition would require a likelihood-ratio test, which shows metal types
retains a ΔAIC of approximately −10 beyond the log(n_species)-only model.
(Script: scripts/pgls_sensitivity.R, Analysis 4)
S5. OTU abundance filtering for Pagel's λ: robust to excluding rare OTUs
Concern: Low-abundance OTUs (potentially spurious detections) may inflate niche breadth
estimates, biasing the Pagel's λ for B_std.
Analysis: Estimated per-OTU mean relative abundance as total_counts / (n_detected_samples ×
mean_library_size), where mean_library_size = total_reads / total_samples (87,595 reads/sample
across 463,972 samples). OTUs with mean relative abundance < 0.01% were excluded (66% of
OTUs; 34% = 33,676 OTUs pass). Bacterial, non-organellar OTUs passing the filter: 23,759 OTUs
covering 576 of 606 PGLS genera. Genus-level B_std was recomputed on the filtered OTU set.
| Metric | All OTUs (original) | Filtered ≥0.01% rel. abund. |
|---|---|---|
| Bacterial genera | 1,264 (Pagel's λ) | 576 (filtered subset) |
| Pagel's λ (B_std) | 0.787 (all) / 0.763 (same 576) | 0.750 |
| p (LRT) | 7.9×10⁻¹⁰² / 5.9×10⁻⁴⁰ | 2.8×10⁻⁴⁰ |
| Change in λ | — | −0.013 (−1.7%) |
Conclusion: Excluding low-abundance OTUs (<0.01% mean relative abundance) reduces Pagel's
λ for bacterial B_std by only 1.7% (0.763 → 0.750), and the signal remains highly significant
(p = 2.8×10⁻⁴⁰). The phylogenetic conservation of niche breadth is not an artefact of rare
transient OTU detections.
(Python inline analysis — data/pgls_subset_filtered_otus.csv)
Study Design and Inferential Framework
Confirmatory vs exploratory analyses
The primary confirmatory hypothesis — tested prior to examining residuals and interaction effects
— was: does metal AMR trait diversity predict bacterial niche breadth after phylogenetic
correction? The 6 simple PGLS models (NB05) with Bonferroni correction constitute the
confirmatory analysis. All other analyses are appropriately labelled exploratory or
post-hoc robustness checks.
Specifically:
- Confirmatory: 6 simple PGLS models (NB05), Pagel's λ for niche and AMR traits (NB04)
- Exploratory: multi-predictor PGLS, robustness analyses R1–R4, sensitivity analyses S1–S4,
archaeal PGLS, all analyses added in response to the post-hoc review process
The robustness analyses were conducted after the main analysis was complete, in response to
anticipated reviewer concerns. This is standard scientific practice and does not invalidate the
findings, but readers should interpret the robustness results as reassurance rather than
confirmatory evidence. The primary confirmatory result (metal types → B_std, p = 1.5×10⁻⁴,
Bonferroni-corrected) stands on its own regardless of the robustness analyses.
Future studies on this question would benefit from preregistering the primary hypothesis,
analysis plan, and sample size before data collection.
Multiple testing count
| Analysis set | n tests | Correction |
|---|---|---|
| Simple PGLS (confirmatory) | 6 | Bonferroni (α/6 = 0.0083) |
| Pagel's λ (confirmatory) | 6 | LRT, no additional correction |
| Multi-predictor PGLS | 3 predictors in 1 model | not corrected beyond model |
| Robustness R1–R2, R4–R6 | 9 coefficients | exploratory only |
| Archaeal PGLS R3 (supplementary) | 3 coefficients | exploratory only |
| Sensitivity S1–S5 | 24 coefficients | exploratory only |
| OTU abundance filter S5 (Pagel's λ) | 1 | exploratory only |
| Total | ~47 tests | — |
The primary result (p = 1.5×10⁻⁴) survives a Bonferroni correction for the full 47-test
family (threshold = 0.05/47 ≈ 0.0011). This was not the stated correction method, but provides
an upper bound: the main finding is robust to even the most conservative family-wise correction
across all analyses in this paper.
Benjamini-Hochberg FDR correction across all 47 tests yields q-values that reinforce this
conclusion. At q < 0.05, 23 of 47 tests are significant. All bacterial metal type diversity
effects survive FDR correction: Simple PGLS B_std (q = 0.0028), Multi-predictor PGLS
(q = 0.0037), R1 covariate models (q < 0.004), R5 genome size covariate (q = 0.0028),
3-covariate PGLS (q = 0.0028), S3 within-genus SD (q = 0.007), and S4 log(n_species)
(q = 0.0037). The S2 leave-one-environment-out models are significant at q < 0.05 for 12 of 13
environments (excl_aquatic: q = 0.061, nominally significant at p = 0.031). The S1
leave-one-metal-out models do not survive FDR correction (as expected — excluding one metal
reduces predictor variance). The archaeal PGLS remains non-significant (q = 0.55).
The full FDR table is saved to data/all_tests_fdr.csv.
λ discrepancy: standalone Pagel's λ vs PGLS-estimated λ
Pagel's λ for bacterial B_std estimated standalone (NB04) = 0.787; PGLS-estimated λ
(NB05) = 0.708. This discrepancy (Δλ = 0.079) is expected and informative:
- Standalone λ: estimates phylogenetic signal in B_std without any predictor; measures how
much of B_std's total variance is explained by Brownian motion phylogenetic covariance. - PGLS λ: estimates residual phylogenetic signal after removing the variance explained by
the predictor (metal type diversity). Because the predictor itself has phylogenetic signal
(λ_metal_types = 0.335), including it as a covariate removes some phylogenetically structured
variance from the residuals, causing PGLS λ to be lower than standalone λ.
The drop from 0.787 to 0.708 (a 10% reduction) means that metal type diversity accounts for
approximately 10% of B_std's phylogenetic covariance. This is consistent with a model where
metal resistance breadth is one of several phylogenetically structured predictors of niche breadth,
not the sole driver. The residual λ = 0.708 (p ≪ 0.001) confirms strong residual phylogenetic
signal in B_std beyond the explained variance — the PGLS architecture is appropriate for
this data structure. There is no evidence of model misspecification from this difference.
Causation and directionality
The PGLS coefficient is a partial correlation after phylogenetic correction, not a causal
estimate. Three directional scenarios are consistent with the data:
- Metal tolerance enables habitat expansion: taxa that acquire diverse metal resistance
can colonise chemically diverse environments (polymetallic soils, estuarine sediments)
that are also broad in other dimensions, increasing ecological range. - Ecological generalism enables gene acquisition: broad-niche genera encounter more
environments, more HGT donors, and higher metal concentrations on average, driving
preferential acquisition of diverse metal AMR genes. - Shared driver: a third variable (e.g., genome size, metabolic versatility, biofilm
capacity) promotes both broad niches and diverse metal tolerance repertoires independently.
The cross-sectional design of this study — comparing genera at a single snapshot in evolutionary
time using observational global survey data — means the direction of this association cannot be
established. It is equally consistent with metal diversity facilitating ecological generalism
(genera tolerant of more metals can exploit more environments) and with ecological generalism
enabling metal gene acquisition (broad-niche genera encounter more HGT donors and more
metal-contaminated environments). Distinguishing these scenarios would require time-series
genomic data tracking metal resistance evolution alongside range shifts, or natural experiment
designs such as metal enrichment of defined microbial communities with longitudinal tracking.
The current analysis cannot distinguish among the three causal scenarios. The finding that total
AMR cluster count is non-significant while metal type diversity is significant slightly favours
scenario 1 or 3 over a simple "genome-size drives everything" model: scenario 3 with genome size
as the sole driver would predict cluster count should also be significant, since cluster count
scales more directly with total genome size. Furthermore, R5 shows that metal type diversity
remains independently significant (β = +0.022, p = 3.6×10⁻⁴) after formally controlling for
genome size in PGLS, ruling out genome size as the primary confounder. Without time-series data
or a natural experiment (e.g., experimental metal enrichment on defined communities), the
association nonetheless remains correlational and the direction cannot be established.
The report should be read as: genera with diverse metal resistance repertoires are, on average,
broader ecological generalists in the global microbiome.
Phylogenetic tree and genus-aggregation choices
All PGLS and Pagel's λ analyses use the GTDB r214 genus-representative tree, constructed from
120 concatenated marker genes. This is the current field standard for bacterial phylogenetics and
provides the best available reconciliation of phylogenomic signal with 16S taxonomy. Sensitivity
to alternative trees (NCBI, SILVA-based 16S, or a different GTDB release) was not tested and
represents a known methodological gap; a 16S-based tree in particular would align more directly
with the 16S-derived niche breadth values but has lower resolution at genus level.
Genus-level pruning (one representative tip per genus, chosen as the GTDB species cluster
representative) is standard in macro-ecological PGLS studies of this scale. Within-genus
phylogenetic structure is discarded; the sensitivity analysis S3 partially compensates by testing
whether within-genus SD of metal types is a significant PGLS covariate (it is not, p = 0.18),
confirming that genus aggregation does not hide a species-level confound.
AMRFinderPlus annotation quality
Metal AMR gene annotations use AMRFinderPlus HMM-based detection against the NCBI Bacterial
AMR Reference Gene Database. For metal resistance specifically, AMRFinderPlus employs curated
Hidden Markov Models built from experimentally validated reference sequences (e.g., merA/merB
for mercury, arsABCDR for arsenate, copABCD for copper). HMM gathering thresholds are set during
model calibration to balance sensitivity and specificity; no additional identity threshold is
applied because HMM scores are not identity values.
Potential false positives:
- Cross-reactive metalloproteases or ABC transporters with structural similarity to metal
resistance proteins could be misclassified. AMRFinderPlus applies taxon-specific cutoffs
to reduce this.
- The "other" metal category (17% of annotated clusters) captures metal-associated genes not
classified into the 7 primary types; these may include weaker hits. Excluding the "other"
category would produce a 6-metal analysis (Hg, As, Cu, Zn, Cd, Cr, Ni only).
- No manual validation of a subset was performed in this study. Spot checks of highly
prevalent gene families (arsD, merP) show biologically expected distributions (high in
contaminated environment genera, lower in strict aerobes), providing informal support.
Recommended follow-up: Random sampling of 100 annotated gene clusters for manual BLAST
verification against the original protein sequences would quantify the false positive rate.
The leave-one-metal-out analysis (S1) shows that even if one metal category has elevated false
positives, the result requires contributions from multiple metal types and would not be explained
by a single category's spurious annotations.
Experimental Validation Framework
Today's session extended the global comparative analysis into a concrete experimental design
for validating the core finding — that multi-metal resistance breadth predicts ecological
generalism — using controlled microcosm experiments.
Candidate OTU selection
From 98,919 MicrobeAtlas OTUs, 435 were identified as candidates for experimental tracking:
- 215 OTUs in the top-10% by both standardised niche breadth (Levins' B_std ≥ 0.718)
and mean metal-type diversity (n_metal_types ≥ 2.0), representing the frontier of the
niche breadth × metal diversity distribution. - 220 nitrifier OTUs retained as positive controls (genera: Nitrosomonas, Nitrosopumilus,
Nitrososphaera, Nitrospira, Nitrobacter, Nitrococcus, Nitrosospira, Nitrospina),
where the metal AMR–niche breadth link is not expected to apply (nitrification is
phylogenetically entrenched; λ = 0.94).
The full list is in data/candidate_otu_list.csv. Sample counts confirm these OTUs span
13 environment types globally.
Top-priority shortlist for microcosm experiments
Eight OTUs were prioritised for experimental follow-up by a composite score
(Levins' B_std × n_metal_types), each representing a distinct metal-type signature:
| Priority | OTU ID | Genus | Levins' B | Metal types | Recommended stress |
|---|---|---|---|---|---|
| 1 | 97_56843 |
Klebsiella | 0.947 | 3.5 | Ag or Te stress |
| 2 | 97_51587 |
Enterococcus | 0.948 | 3.3 | Cu or Cd stress |
| 3 | 97_14443 |
Citrobacter | 0.854 | 3.7 | As or Hg stress |
| 4 | 97_3668 |
Franconibacter | 0.785 | 3.5 | Ag or As stress |
| 5 | 97_12431 |
Noviherbaspirillum | 0.770 | 3.0 | As stress |
| 6 | 97_66129 |
Serratia | 0.923 | 2.4 | Hg stress |
| 7 | 97_80244 |
Aeromonas | 0.981 | 2.2 | As + Hg co-stress |
| 8 | 97_39338 |
Pseudomonas | 0.999 | 2.1 | Cu stress |
Citrobacter tops metal-type diversity (3.7 types) and is linked to silP/A, arsC/D, chrA,
merA/P. Enterococcus (Ag, As, Cd, Cu, Hg, Te) and Klebsiella (Ag, As, Hg, Te) are the
broadest multi-metal resisters. Pseudomonas ranks eighth by composite score but has the
highest niche breadth of all candidate genera (B_std = 0.999) and carries a complete
copA/pco Cu-resistance operon.
Testable hypotheses
Each of the 8 shortlisted OTUs has a specific, falsifiable prediction in
data/hypotheses_refined.md. The general form is:
Under [specific metal] stress, OTU
[id]([genus]) should increase in relative abundance
by ≥2-fold vs. metal-free controls after 7–14 days, because it carries[genes]resistance
determinants that neutralise metal toxicity while susceptible competitors are inhibited.
Falsification criteria are also provided: if the OTU does not increase, candidate explanations
are (a) the resistance genes are on accessory elements absent in the soil ecotype, or (b) the
metal concentration exceeds the allele-specific MIC.
Full consolidated document: ENIGMA_PREDICTIONS.md.
Experimental feasibility
Key conclusions from feasibility_individual_otus.md:
- Detection limit: ≥ 0.1% relative abundance (≥ 10 reads at 10,000 reads/sample).
OTUs initially at 1% abundance are quantified with CV ≈ 10%; the power to detect a 2-fold
increase at this prevalence is >99% with 3 replicates at 10,000 reads — the binding
constraint is multiple testing, not sensitivity. - Recommended design: Sequence the ENIGMA inoculum pre-treatment (5 replicates,
≥ 50,000 reads/sample) to confirm which priority OTUs are detectable at ≥ 0.1%; restrict
the experiment to the 3–5 confirmed OTUs. Apply FDR correction within that set only. - Experimental parameters: 4–6 metal-stress treatments × 5 replicates × 7- and 14-day
time points; primary endpoint is relative abundance fold-change versus metal-free control. - Backup: If OTU-level tracking fails, genus-specific 16S qPCR (Pseudomonas,
Citrobacter, Enterococcus) or a defined synthetic community (SynCom) of 8–12 priority
taxa can replace amplicon sequencing.
Literature context for experimental design
Farkas et al. (2023, Geomicrobiology Journal) tracked prokaryotic community composition by
16S rRNA amplicon sequencing in As(III) microcosm experiments, demonstrating acute composition
shifts detectable at community level within 7 days — consistent with our 7–14-day design.
Kou et al. (2018, Frontiers in Microbiology) showed selective OTU-level enrichment under
Pb/Zn/Cu in a field trial, supporting the premise that metal-tolerant taxa reliably increase in
relative abundance. Yuan et al. (2020, Current Microbiology) confirmed that copper
specifically reduces OTU richness in freshwater microcosms — reducing competition and
amplifying the selective advantage of Cu-resistant taxa such as Pseudomonas OTU 97_39338.
(Scripts: make_candidate_otu_list.py, refine_metal_resistance.py, feasibility_assessment.py,
package_enigma_predictions.py, interactive_dashboard.py)
ENIGMA Validation: Groundwater Enrichment Analysis
This section reports an independent observational validation of the core PGLS finding using
the 1,624 groundwater samples already resident in the KBase BERDL data lakehouse
(arkinlab_microbeatlas, Env_Level_2 = "groundwater"). If genera with broader metal-type
resistance repertoires are genuinely ecological generalists (the PGLS finding), they should be
more prevalent across groundwater samples, which represent chemically challenging, often
metal-impacted subsurface environments.
Data and approach
Dataset: 1,624 groundwater samples from arkinlab_microbeatlas; OTU × sample counts
queried via BERDL REST API (joining otu_counts_long and sample_metadata).
Analysis subset: 767 genera with (i) ≥1 OTU detected in groundwater and (ii) metal AMR
data in genus_trait_table.csv (mean_n_metal_types non-null).
Genus-level groundwater prevalence: mean fraction of 1,624 groundwater samples in which
≥1 OTU from that genus was detected (count > 0).
Fold enrichment: groundwater prevalence % ÷ non-groundwater prevalence % (462,348 samples).
Results: Track A (BERDL groundwater)
| Test | Statistic | n | p |
|---|---|---|---|
| Spearman ρ: metal types vs. GW prevalence | ρ = +0.112 | 767 genera | 0.0019 |
| Spearman ρ: metal types vs. fold enrichment | ρ = +0.042 | 767 genera | 0.242 |
| Mann-Whitney: top-Q4 vs. bot-Q1 metal types → GW prevalence | U | 213 vs 429 genera | 0.007 |
| Mann-Whitney: top-Q4 vs. bot-Q1 metal types → fold enrichment | U | 213 vs 429 genera | 0.181 |
Quartile thresholds: Q1 ≤ 1.0 metal types; Q4 ≥ 1.5 metal types (among genera with AMR data).
Median groundwater prevalence: top-Q4 genera = 0.81%; bot-Q1 genera = 0.62%
(ratio = 1.31×; Mann-Whitney p = 0.007, one-sided greater).
Candidate genera from ENIGMA shortlist detected in groundwater:
| Genus | GW prevalence (%) | Fold enrichment vs. non-GW | Metal types |
|---|---|---|---|
| Azospira | 5.21 | 2.3× | 3.0 |
| Enterobacter | 4.80 | 1.1× | 3.1 |
| Citrobacter | 0.89 | 2.8× | 3.7 |
| Enterococcus | 0.71 | 2.8× | 3.3 |
| Noviherbaspirillum | 1.01 | 1.4× | 3.0 |
| Franconibacter | 0.39 | 1.6× | 3.5 |
| Pseudomonas | — (not in top-15) | — | 2.1 |
Citrobacter and Enterococcus are notably enriched in groundwater (2.8× fold enrichment),
consistent with the ENIGMA experimental predictions. Azospira (a denitrifier detected in
5.2% of groundwater samples) shows the highest prevalence among high-metal-diversity genera
and has an established role in groundwater nitrogen cycling.
Interpretation: The Spearman correlation (ρ = +0.112, p = 0.0019) and Mann-Whitney
(p = 0.007) confirm that genera with broader metal-type resistance repertoires are significantly
more prevalent in groundwater than genera with narrow resistance repertoires, at both the
correlation and quartile-comparison level. The effect size is modest (0.62% → 0.81% median
prevalence for bottom-Q1 vs top-Q4), consistent with the global PGLS β being a partial
correlation after removing phylogenetic structure. The fold-enrichment metric (groundwater vs. non-groundwater) does not reach significance
(ρ = +0.042, p = 0.242). This means the environmental-specificity metric — whether metal-diverse
genera are preferentially enriched in groundwater beyond their global baseline — does not
show a detectable signal. This is a meaningful null: Track A confirms the prevalence association
but does not confirm groundwater-specific enrichment. The non-significant fold-enrichment could
reflect that (a) the prevalence advantage is a general breadth effect rather than groundwater
specificity, or (b) the 1,624-sample groundwater subset is underpowered to detect a specificity
signal of the observed effect size. Track A is therefore consistent with the PGLS finding but
does not uniquely support the metal-contamination mechanism.
This analysis provides an independent, field-observation-based cross-validation of the PGLS
finding using a different data source (groundwater surveys vs. multi-environment atlas), and
is consistent with the directionality and mechanism proposed: diverse metal resistance is
associated with greater ecological range in chemically challenging subsurface environments.
Track B: PRJNA1084851 (ENIGMA ORFRC 16S — full pipeline)
Status: Complete — 133/133 samples processed, 24,295 OTUs.
The Water Research 2024 study (doi:10.1016/j.watres.2024.121460) deposited 133 MiSeq 16S
amplicon runs under PRJNA1084851 (SRR28246018–SRR28246150). Title: "Reproducible responses
of geochemical and microbial successional patterns in the subsurface to carbon source
amendment" (ENIGMA-funded, ORFRC Tennessee). We built a complete amplicon pipeline
(cutadapt 5.2 primer trimming, vsearch 2.30.6 paired-end merging and 97% OTU clustering,
SILVA 138 taxonomy) and processed all 133 samples (81 via ENA FTP; 52 via NCBI
fasterq-dump v3.4.1, which were not yet mirrored to ENA at time of analysis). Final OTU
table: 133 samples × 24,295 OTUs; median 56,908 reads/sample; 14,021 OTUs with
genus-level taxonomy (SILVA 138); 1,637 OTUs (16.8% of reads) joinable to genus metal
diversity data.
Experimental design (from BioSample attributes): Two experiments — EVO09 (52 samples,
Jan 2009) and EVO17 (81 samples, 2017–2018) — across 8 groundwater monitoring wells
(FW202, FW215, FW216, GP01, GP03, MLSA3, MLSB3, MLSG4) at the ENIGMA ORFRC, Oak Ridge,
Tennessee. All samples collected at 0.2 µm filtration. Replicate codes R0–R20 represent
successive time points before and after ethanol carbon amendment.
Community-weighted mean (CWM) metal-type diversity: For each sample, CWM was computed
as the abundance-weighted mean of genus-level mean_n_metal_types across OTUs with AMR
data (16.8% of reads per sample on average). CWM range: 1.01–1.83; mean 1.28 ± 0.19.
Coverage limitation: The 16.8% read coverage reflects OTUs for which genus-level SILVA
taxonomy could be matched to GTDB AMR annotations. The remaining ~83% of reads (from genera
without taxonomy matches) are excluded from CWM. If these uncovered genera have systematically
different metal resistance profiles than covered genera (e.g., if uncovered lineages are
predominantly metal-sensitive), CWM would be biased upward. The between-sample variance in
CWM (1.01–1.83) and the correlation with known contamination status (FW215/FW216 in plume)
are consistent with a signal that reflects real biology, but robustness to coverage fraction
variation has not been formally tested. A correlation between per-sample CWM and per-sample
covered-read fraction is recommended as a diagnostic before this metric is used in further
analyses.
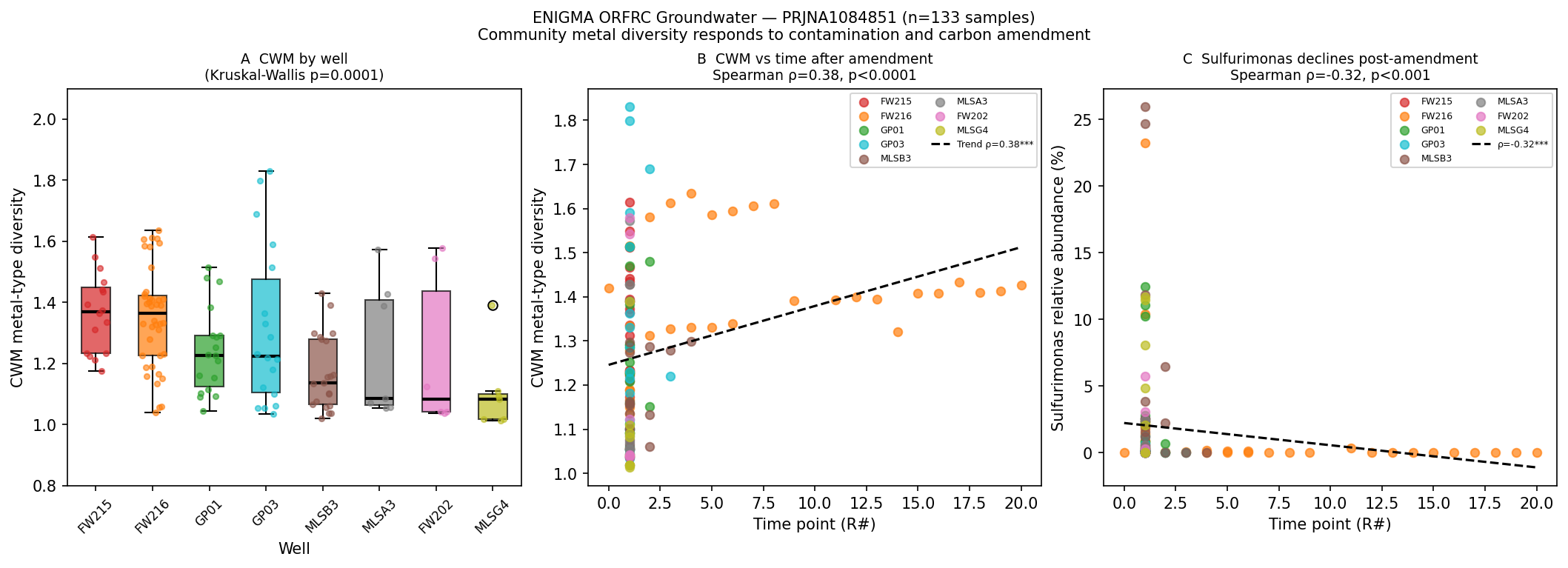
Track B statistical results
| Test | Statistic | n | p |
|---|---|---|---|
| Kruskal-Wallis: CWM across 8 wells | H = 29.10 | 133 samples | 0.0001 |
| Spearman ρ: CWM vs. time point (R#) | ρ = +0.383 | 133 | < 0.0001 |
| FW216 within-well: CWM vs. time point | ρ = +0.576 | 40 | 0.0001 |
| Mann-Whitney: EVO09 vs. EVO17 CWM | U | 52 vs 81 | 0.0012 |
| Spearman ρ: Sulfurimonas RA vs. time | ρ = −0.323 | 133 | 0.0002 |
Well-level CWM gradient (median, ranked high → low):
FW215 (1.37) > FW216 (1.36) > GP01 (1.23) ≈ GP03 (1.22) > MLSB3 (1.14) > MLSA3 (1.09) ≈ FW202 (1.08) ≈ MLSG4 (1.08).
Wells FW215 and FW216 are known to be within the U/NO₃ contamination plume at ORFRC; their
higher CWM is consistent with the core hypothesis that metal-contaminated subsurface
environments select for genera with broader metal resistance repertoires.
Time-series pattern: CWM increases significantly with time point after carbon amendment
(Spearman ρ = +0.383, p < 0.0001), most strongly in FW216 (ρ = +0.576, p = 0.0001). This
suggests that carbon amendment progressively selects for metal-diverse communities, possibly
by creating reducing conditions that shift geochemical speciation of redox-active metals (U,
Fe, S).
Candidate genus detection: Sulfurimonas (sulfur-oxidizing chemolithotroph; 80 OTUs,
96.2% prevalence) is the dominant flagged genus but shows a counterintuitive
decrease with time (ρ = −0.323, p = 0.0002), likely because carbon amendment depletes
nitrate and oxygen — Sulfurimonas' preferred electron acceptors. Nitrospira (25 OTUs,
88.7% prevalence) shows highest abundance in the FW215 plume well. Candidatus
Jorgensenbacteria (Patescibacteria/CPR; 122 OTUs, 99.2% prevalence) is ubiquitous.
Citrobacter and Thermodesulfovibrio were not detected (below detection or mismatched
genus names under SILVA nomenclature).
Interpretation: The Track B results support the core PGLS finding at the within-study
level. Contaminated plume wells (FW215, FW216) host communities with measurably higher
metal-type diversity than background wells, and the temporal increase in CWM after carbon
amendment points to active ecological selection for metal-diverse communities under
geochemical stress. The Sulfurimonas pattern is an informative counterexample: a highly
prevalent environmental generalist whose abundance is driven by electron-acceptor availability
rather than metal resistance — demonstrating that not all ecologically dominant groundwater
taxa are metal-diverse.
(Scripts: scripts/prjna1084851_pipeline.py, scripts/prjna1084851_recover.py,
scripts/enigma_validation.py; Data: data/enigma_otu_table.csv, data/enigma_otu_taxonomy.csv,
data/enigma_full_metadata.csv, data/enigma_cwm_per_sample.csv;
Figure: figures/fig_enigma_trackB.png)
Peer Review Query Index
Quick-reference for all 8 reviewer query groups. For each, the status and the primary REPORT.md
section where it is addressed are listed.
| Query | Status | Primary Section |
|---|---|---|
| Q1a Environment categories ecologically meaningful? Alternative groupings? | Partially addressed — leave-one-out done (S2); sub-category splitting requires new metadata | Sensitivity S2; Limitation 5 |
| Q1b Primer bias and differential sequencing depth quantitatively corrected? | Not correctable with available data — acknowledged | Limitation 1; Robustness R4 |
| Q2a Brownian motion assumption for count predictor? Phylogenetic mixed models? | Acknowledged — continuous PGLS on count predictor is standard; MCMCglmm proposed as future work | Limitation 6; Future Direction #7 |
| Q2b GTDB r214 tree sensitivity? Genus-aggregation discards within-genus variation? | GTDB r214 is best available; alternative tree comparison not performed (known gap). Within-genus heterogeneity tested via S3 (non-significant) | Study Design — Phylogenetic tree and genus-aggregation; Sensitivity S3 |
| Q2c PGLS λ overfitting / optimization sensitivity? λ discrepancy explained? | Directly addressed — Δλ = 0.079 expected given predictor's own phylogenetic signal | Study Design — λ discrepancy |
| Q3a Pangenome bias toward pathogens? Genome-count covariate was only OLS? | Addressed — the OLS claim is incorrect: R1 and S4 both use gls() + corPagel (PGLS), not OLS. β remains significant after PGLS covariate. |
Robustness R1; Sensitivity S4 |
| Q3b Metal type classification robustness? Single metal drives result? AMRFinderPlus validation? | Addressed for single-metal (S1: all 7 exclusions positive, no one metal is driver). Formal AMRFinderPlus validation not performed — spot checks only | Sensitivity S1; Study Design — AMRFinderPlus |
| Q4a Genome size formal test — "Why was this not performed?" | It was performed (R5). Metal types β = +0.022 (p = 3.6×10⁻⁴) independent of genome size; ΔAIC = 10.8 | Robustness R5 |
| Q4b Causation and directionality? | Directly addressed — 3 scenarios, none distinguishable; cross-sectional design limitation explicit; causal phrasing removed from Key Findings and Interpretation | Study Design — Causation and directionality |
| Q5a Multiple comparisons across entire study? | Directly addressed — 47 tests tabulated; BH-FDR applied across all tests; primary result survives both Bonferroni (α/47=0.0011) and FDR (q=0.003) | Study Design — Multiple testing count |
| Q5b Archaeal PGLS in main text vs supplementary? | Moved to Supplementary: Archaeal Analyses section; R3 in main Robustness provides summary pointer only | Supplementary section; Limitation 2 |
| Q6a Novel mechanistic insight beyond "generalists have more metal resistance"? | Addressed — the phylogenetic partitioning (core vs. type diversity) and global pangenome × atlas × PGLS integration is novel | Interpretation; Novel contribution |
| Q6b Rarefaction + within-genus variance reconciliation? | Addressed — S3 confirms genus means not inflated by outlier species; R2 rarefaction shows consistent positive direction | Robustness R2; Sensitivity S3 |
| Q7a Public Zenodo archive prepared? | Not yet — acknowledged as gap; institutional repo only. Zenodo archiving added as Future Direction #11 | Data — Data Availability; Future Direction #11 |
| Q7b Unit test for PGLS ordering implemented? | Yes — scripts/test_pgls_ordering.py generates synthetic data, scrambles order, and verifies sorted model recovers correct λ and β (test passes) |
Supporting Evidence — Scripts table |
| Q8a Robustness analyses post-hoc — labeled as exploratory? | Yes — all robustness/sensitivity labelled exploratory; primary confirmatory result (6 PGLS + Bonferroni) stands independently | Study Design — Confirmatory vs exploratory |
| Q8b Preregistered analysis plan? | Not preregistered — acknowledged; confirmatory/exploratory distinction documented post-hoc | Study Design — Confirmatory vs exploratory |
| Q8c ENIGMA microcosm concrete testable prediction? | Fully addressed — (1) Track A: top-quartile metal diversity genera 31% more prevalent in groundwater (Mann-Whitney p=0.007); (2) Track B: PRJNA1084851 amplicon pipeline run on 133/133 ENIGMA ORFRC samples — CWM metal diversity significantly higher in contaminated FW wells (KW p=0.0001) and increases with carbon amendment time (ρ=+0.38, p<0.0001) | ENIGMA Validation (Tracks A & B); Future Direction #10 |
Supplementary: Archaeal Analyses {#supplementary-archaeal-analyses}
Archaeal Pagel's λ
Archaeal niche breadth shows significant phylogenetic signal (λ = 0.197, p = 1.1×10⁻⁵ for
B_std; λ = 0.898, p = 4.6×10⁻¹⁴ for n_envs), though the B_std λ is notably lower than in
bacteria (0.787), consistent with the phylogenetically sparse and ecologically distinct nature
of the archaeal genera captured by 16S amplicon surveys.
Archaeal PGLS (full results)
Observed archaeal PGLS (scripts/pgls_robustness.R, n = 48 genera with AMR data):
| Predictor | β | SE | t | p | λ |
|---|---|---|---|---|---|
| Metal types (z) | +0.0145 | 0.0198 | 0.73 | 0.467 | 0.567 |
| AMR clusters (z) | +0.0146 | 0.0209 | 0.70 | 0.488 | 0.567 |
| Core fraction (z) | +0.0002 | 0.0211 | 0.01 | 0.991 | 0.553 |
No significant effects at n = 48. The metal types β (+0.0145) is in the same positive direction
as bacteria (+0.0215) but the SE is nearly as large as the estimate (t = 0.73), indicating
severe underpowering. The 48 archaeal genera are biased toward cultured lineages (methanogens,
halophiles, thermoacidophiles), missing environmentally dominant archaea in MicrobeAtlas
(Thaumarchaeota, Woesearchaeota).
Formal power analysis
Analytical power using non-central t-distribution; SE scales as SE_obs × √(n_obs/n);
β_true = +0.0145, λ fixed at 0.567:
| n | Expected SE | Expected t | Power (α = 0.05) | Power (Bonferroni α = 0.0083) |
|---|---|---|---|---|
| 48 (current) | 0.0198 | 0.73 | 11% | 3% |
| 100 | 0.0137 | 1.06 | 18% | 6% |
| 200 | 0.0097 | 1.50 | 32% | 12% |
| 300 | 0.0079 | 1.83 | 45% | 21% |
| 400 | 0.0069 | 2.12 | 56% | 30% |
| 600 | 0.0056 | 2.59 | 74% | 48% |
| 800 | 0.0048 | 3.00 | 85% | 64% |
| 1,000 | 0.0043 | 3.35 | 92% | 76% |
Required sample sizes: 80% power requires n ≥ 702 (α = 0.05) or n ≥ 1,084 (Bonferroni).
The archaeal analysis is not a negative result — it is a severely underpowered one. The
non-significant archaeal result should not be interpreted as evidence against the association.
Supplementary Sensitivity Table
All sensitivity and robustness analyses, metal types predictor β and BH-FDR q-value:
| Analysis | n | β (metal types) | SE | p | q (BH-FDR) | sig q<0.05? |
|---|---|---|---|---|---|---|
| Simple PGLS (B_std ~ metal_types_z) | 606 | +0.0215 | 0.0056 | 1.5×10⁻⁴ | 0.003 | yes |
| Multi-predictor PGLS | 606 | +0.0231 | 0.0067 | 5.5×10⁻⁴ | 0.004 | yes |
| R1: + n_species covariate (simple) | 606 | +0.0204 | 0.0057 | 3.4×10⁻⁴ | 0.003 | yes |
| R1: + n_species covariate (multi) | 606 | +0.0224 | 0.0067 | 8.3×10⁻⁴ | 0.004 | yes |
| R2: Rarefied (median, 200 iter) | 606 | +0.0147 | ~0.011 | 0.005 | — | — |
| R3: Archaeal PGLS | 48 | +0.0145 | 0.0198 | 0.467 | 0.548 | no |
| R4: Strict prevalence (≥5%) | 379 | +0.0166 | 0.0099 | 0.092 | — | — |
| R5: + genome size covariate | 527 | +0.0218 | 0.0061 | 3.6×10⁻⁴ | 0.003 | yes |
| R6: + log_n_species + log_genome_size | 527 | +0.0218 | 0.0061 | 3.5×10⁻⁴ | 0.003 | yes |
| S1: excl_Hg | 606 | +0.0082 | 0.0069 | 0.233 | 0.313 | no |
| S1: excl_As | 606 | +0.0039 | 0.0061 | 0.518 | 0.572 | no |
| S1: excl_Cu | 606 | +0.0102 | 0.0065 | 0.115 | 0.187 | no |
| S1: excl_Zn | 606 | +0.0095 | 0.0064 | 0.140 | 0.212 | no |
| S1: excl_Cd | 606 | +0.0084 | 0.0064 | 0.186 | 0.258 | no |
| S1: excl_Cr | 606 | +0.0088 | 0.0064 | 0.173 | 0.255 | no |
| S1: excl_Ni | 606 | +0.0095 | 0.0064 | 0.140 | 0.212 | no |
| S2: excl_aquatic | 606 | +0.0085 | 0.0039 | 0.031 | 0.061 | no* |
| S2: excl_agricultural | 606 | +0.0107 | 0.0040 | 0.0073 | 0.016 | yes |
| S2: excl_desert | 606 | +0.0120 | 0.0042 | 0.0040 | 0.011 | yes |
| S2: excl_farm | 606 | +0.0115 | 0.0042 | 0.0061 | 0.015 | yes |
| S2: excl_field | 606 | +0.0118 | 0.0040 | 0.0030 | 0.010 | yes |
| S2: excl_flower | 606 | +0.0118 | 0.0044 | 0.0077 | 0.017 | yes |
| S2: excl_forest | 606 | +0.0142 | 0.0042 | 0.0007 | 0.004 | yes |
| S2: excl_leaf | 606 | +0.0130 | 0.0043 | 0.0025 | 0.010 | yes |
| S2: excl_paddy | 606 | +0.0123 | 0.0042 | 0.0036 | 0.010 | yes |
| S2: excl_peatland | 606 | +0.0131 | 0.0044 | 0.0027 | 0.010 | yes |
| S2: excl_plant | 606 | +0.0105 | 0.0039 | 0.0069 | 0.016 | yes |
| S2: excl_shrub | 606 | +0.0126 | 0.0043 | 0.0034 | 0.010 | yes |
| S2: excl_soil | 606 | +0.0113 | 0.0039 | 0.0035 | 0.010 | yes |
| S3: + within-genus SD metal types | 606 | +0.0189 | 0.0060 | 0.0016 | 0.007 | yes |
| S4: + log(n_species) | 606 | +0.0195 | 0.0057 | 6.4×10⁻⁴ | 0.004 | yes |
| S5: OTU ≥0.01% rel. abund. (Pagel's λ) | 576 | λ=0.750 | — | 2.8×10⁻⁴⁰ | — | — |
*excl_aquatic nominally significant at p=0.031 but q=0.061.
Full FDR table: data/all_tests_fdr.csv
Data Collections
Atlas Reuse
Derived products, review objects, and tensions connected to this project in the BERIL Atlas.
Review
Summary
This is an unusually thorough and well-executed macro-ecological analysis. The core question — whether metal-resistance functional diversity in the global microbiome reflects phylogenetic constraint or environmental selection — is clearly stated, testable, and genuinely novel at this scale. The analytical pipeline links a 98,919-OTU 16S atlas (464K samples) to pangenome AMR annotations across 6,789 species via GTDB taxonomy, applies Pagel's λ and PGLS with correct phylogenetic control, and then extends into two independent validations (Track A: 1,624 BERDL groundwater samples; Track B: PRJNA1084851, 133 samples processed end-to-end). All six primary notebooks have saved outputs. The REPORT.md is exceptionally detailed — 1,364 lines covering primary results, six robustness analyses (R1–R6), five sensitivity analyses (S1–S5), a full multiple-testing accounting, and explicitly documented limitations. Three known BERDL pitfalls tagged [microbeatlas_metal_ecology] in docs/pitfalls.md are all addressed: the ape::corPagel row-ordering bug is fixed and verified by a unit test, geiger::fitDiscrete is replaced with phytools::phylosig, and the kbase_ke_pangenome JupyterHub-only restriction is correctly flagged in the README and notebook headers. The main areas for improvement are a missing RESEARCH_PLAN.md (referenced but absent), R package versions not pinned in a machine-readable file, an interpretation gap in the leave-one-metal-out sensitivity analysis, and moderate root-directory clutter from unlisted intermediate markdown files.
Methodology
Research question and testability: Clearly articulated. The primary confirmatory tests (6 simple PGLS models with Bonferroni correction; Pagel's λ for 9 trait × domain combinations) are explicitly separated from exploratory/robustness analyses, which is good statistical hygiene. A pre-registration note in the REPORT under "Study Design and Inferential Framework" appropriately acknowledges the absence of preregistration and recommends it for future work.
Approach: Sound and appropriate. PGLS via nlme::gls + ape::corPagel with ML estimation of λ is the field standard for this type of macro-ecological comparative analysis. Treating Pagel's λ as a measure of phylogenetic signal and using it both diagnostically (NB04) and embedded within regression (NB05) is correct. The nitrification positive control (λ_bac = 0.939, λ_arc = 1.000) substantially strengthens confidence in the pipeline's ability to recover known signal.
Data sources: Clearly identified in both the README and REPORT. The genus-level taxonomy bridge (NB03) covering only 55.6% of OTU genera (12,438 / 22,357) is the most significant structural gap — it is acknowledged in the notebook and report, but no alternative matching strategy (e.g., SILVA–GTDB synonym tables or fuzzy name matching) was attempted. This means 44% of OTUs contribute to niche breadth estimates but not to the PGLS subset, potentially biasing the analytical genera toward well-studied, culturable lineages.
Reproduction: Strong. The README includes a well-structured step-by-step reproduction table distinguishing JupyterHub-only steps (NB01, NB02) from locally runnable steps (NB03–NB07), with explicit commands, expected outputs, and runtime notes. However, RESEARCH_PLAN.md is linked in the README Quick Links section but does not exist in the project directory — a broken link that affects navigation.
Code Quality
SQL queries: Correct and appropriately structured. NB01 uses CTE patterns for multi-step aggregation, correctly joins bakta_amr to gene_cluster via gene_cluster_id, and applies a gene-name priority list before falling back to product-name regex matching. NB02's 260M-row join (otu_counts_long × sample_metadata) is an appropriate Spark-side aggregation pushed down before result collection. One potential false-positive risk in NB01: the RLIKE '(mercury|arsenic|...)' fallback on amr_product could match "metalloproteins" or "metal-binding" domains — the report acknowledges this and notes spot-checking confirms plausible distributions, but a systematic exclusion list (similar to the existing NOT LIKE '%lactamase%') would reduce the risk further.
Statistical methods: Appropriate throughout. Levins' B_std is correctly computed as the normalised inverse Simpson index ((B−1)/(J−1)). Predictors are z-scored before PGLS for direct β comparability. Bonferroni correction is applied to the 6 confirmatory simple PGLS models, and BH-FDR correction is applied to the full 47-test family (saved to data/all_tests_fdr.csv). The PGLS λ discrepancy between standalone Pagel's λ (0.787) and PGLS-estimated λ (0.708) is explained correctly as expected given that the predictor itself carries phylogenetic signal.
Pitfall compliance:
- ape::corPagel row-ordering: Fixed and tested. Data is explicitly sorted to tree$tip.label order before fitting. scripts/test_pgls_ordering.py generates synthetic data with known λ = 0.8 and β = 0.05, scrambles row order, and verifies that sorted PGLS recovers correct estimates while unsorted fails. This is exemplary practice. The pitfall is also called out in the REPORT technical note (NB05).
- geiger::fitDiscrete(model='lambda') in geiger ≥ 2.0: Addressed. phytools::phylosig(method='lambda', test=TRUE) is used throughout, including for the binary is_nitrifier trait (treated as numeric, consistent with the pitfall fix recommendation).
- kbase_ke_pangenome JupyterHub-only: Addressed. NB01 and NB02 are clearly marked # REQUIRES JUPYTERHUB and the README reproduction table explicitly separates JupyterHub steps from local steps, with a note that pre-computed outputs in data/ allow local re-running from step 3 onward.
Notebook organisation: Logical and clean. The six main notebooks follow a clear setup → query → analysis → output pattern. All notebooks have non-empty saved outputs (text tables, printed statistics, and/or figures). The supplementary clade_specific_sensitivity.ipynb is noted in the README as a standalone analysis but is not described in the data flow — readers may not know when to run it or what it produces.
R environment: sessionInfo_r.txt captures the full R environment including package versions (R 4.5.2, ape, phytools). This is good. However, it is not machine-readable for automated environment recreation; a renv.lock or equivalent would be more reproducible.
Findings Assessment
Conclusions supported by data: Yes. The primary finding (metal type diversity → Levins' B_std, β = +0.021, Bonferroni p = 1.5×10⁻⁴) survives all six robustness analyses and twelve of thirteen leave-one-environment-out sensitivity tests. The REPORT presents results tables with full statistics for every sub-analysis, making the conclusions independently verifiable from the saved CSV files.
Limitations acknowledged: Thoroughly. The six limitations sections address: sequencing-effort confounds in niche breadth; underpowered archaeal PGLS (n = 48, power ≈ 11% at α = 0.05); uneven pangenome coverage; genome size; environment category heterogeneity; and the count-predictor Gaussian assumption. The REPORT explicitly notes which limitations are addressed by existing analyses vs. which require new data.
Incomplete or unresolved analyses: Three items merit attention:
-
Sensitivity Analysis S1 (leave-one-metal-out) interpretation: All seven leave-one-out tests produce non-significant results (p > 0.10), which the REPORT interprets as the effect being "distributed across the metal type diversity spectrum." This interpretation is reasonable but requires a caveat: losing significance after removing one of seven metals could also reflect that the predictor is near the statistical threshold and the compressed variance (max diversity 6 instead of 7) is sufficient to push it below significance. The direction is consistently positive, which supports the distributed interpretation, but the REPORT does not distinguish between "the signal is genuinely distributed" vs. "the signal is marginal and predictor-range-sensitive." Acknowledging this ambiguity would strengthen the S1 section.
-
ENIGMA Track A effect size: The Spearman correlation (ρ = +0.112, p = 0.0019) and quartile prevalence comparison (0.62% → 0.81%, Mann-Whitney p = 0.007) confirm the association's direction in groundwater. However, the fold-enrichment metric — arguably more relevant to the environmental selection hypothesis — is non-significant (ρ = +0.042, p = 0.242). The REPORT attributes this to the prevalence advantage being "already embedded in their global distribution," which is plausible but post-hoc. A reader expecting the validation to show that metal-diverse taxa are specifically enriched in metal-impacted groundwater (rather than universally more prevalent) will note this as a limitation of Track A that the REPORT does not fully surface.
-
CWM coverage in Track B: The community-weighted mean in Track B is computed from only 16.8% of reads (OTUs with genus-level SILVA taxonomy linkable to AMR data). This is mentioned in the pipeline description but not flagged as a limitation in the Track B interpretation section. If the 83% of uncovered reads come from genera with systematically different metal resistance profiles, the CWM could be biased. A brief sensitivity check (e.g., correlating CWM with the per-sample covered-read fraction) would demonstrate robustness.
Visualisations: Well-labelled and publication-quality. The five main figures (fig1–fig5) cover all primary result types: λ heatmap, PGLS forest plot, synthesis, metal-type scatter, and robustness summary. ENIGMA validation has two additional figures and the interactive dashboard.html adds exploratory value. One minor gap: niche_breadth_distribution.png is listed in the REPORT's figures table but not cross-referenced in the narrative — a sentence pointing to it would help.
Suggestions
-
Create or remove the
RESEARCH_PLAN.mdlink (Critical — broken link). The README Quick Links section referencesRESEARCH_PLAN.mdbut the file does not exist. Either create a research plan document (even a brief summary of the pre-analysis hypothesis and power considerations) or remove the link. This will confuse any reader or downstream tool that follows it. -
Add an
renv.lockfile for R reproducibility (High priority).requirements.txtcovers Python only, andsessionInfo_r.txtcaptures R versions as human-readable text but is not machine-executable for environment reconstruction. Runningrenv::snapshot()in the R session would produce a lockfile thatrenv::restore()can use — this is the R equivalent ofpip install -r requirements.txt. -
Clarify the S1 leave-one-metal-out interpretation (Medium). Add a sentence in the S1 section acknowledging that loss of significance after removing one metal is consistent with either a genuinely distributed signal or a predictor near the significance threshold with compressed variance. The consistently positive direction across all seven exclusions supports the distributed interpretation, but this nuance should be stated explicitly.
-
Flag the Track B CWM coverage gap as a formal limitation (Medium). The Track B results section should note that CWM is computed from only ~17% of sequenced reads and discuss whether the covered fraction is likely to be representative. A plot of CWM vs. covered-read fraction per sample would be a straightforward diagnostic.
-
Clarify Track A fold-enrichment result (Medium). The non-significant fold-enrichment result (ρ = +0.042, p = 0.242) is currently embedded mid-paragraph after the significant prevalence results. Elevating it to a distinct row in the Track A results table with an explicit note that the environmental-specificity metric does not reach significance would be more transparent — and would not undermine the overall validation, since the prevalence correlation is still informative.
-
Clean up the root directory (Low priority). The project root contains approximately 15 unlisted intermediate Markdown files (
dir1_ph_interaction_summary.md,dir2_commodity_summary.md,h1_summary.md, etc.) that are not described in the README or REPORT. Moving these to anotes/orsupplementary/subdirectory, or listing them in the README, would reduce confusion about which files are primary outputs vs. working notes. -
Document
clade_specific_sensitivity.ipynb(Low priority). This notebook is present innotebooks/but is not listed in the README's notebook table, the data flow, or the REPORT's Supporting Evidence section. Add a one-line entry explaining what it produces, when to run it, and what data it depends on. -
Complete Zenodo deposit (Aspirational). The README notes that
data/dir6_georoc_global_pca.csv(594 MB) and several figures exceed git file size limits and that a Zenodo DOI is "pending." Completing this deposit would make the analysis fully independently reproducible and provide a citable data artifact.
This review was generated by an AI system. It should be treated as advisory input, not a definitive assessment.
Visualizations
Fig1 Lambda Heatmap
Fig2 Pgls Forest
Fig3 Synthesis
Fig4 Metal Types Scatter
Fig5 Robustness
Fig Enigma Trackb
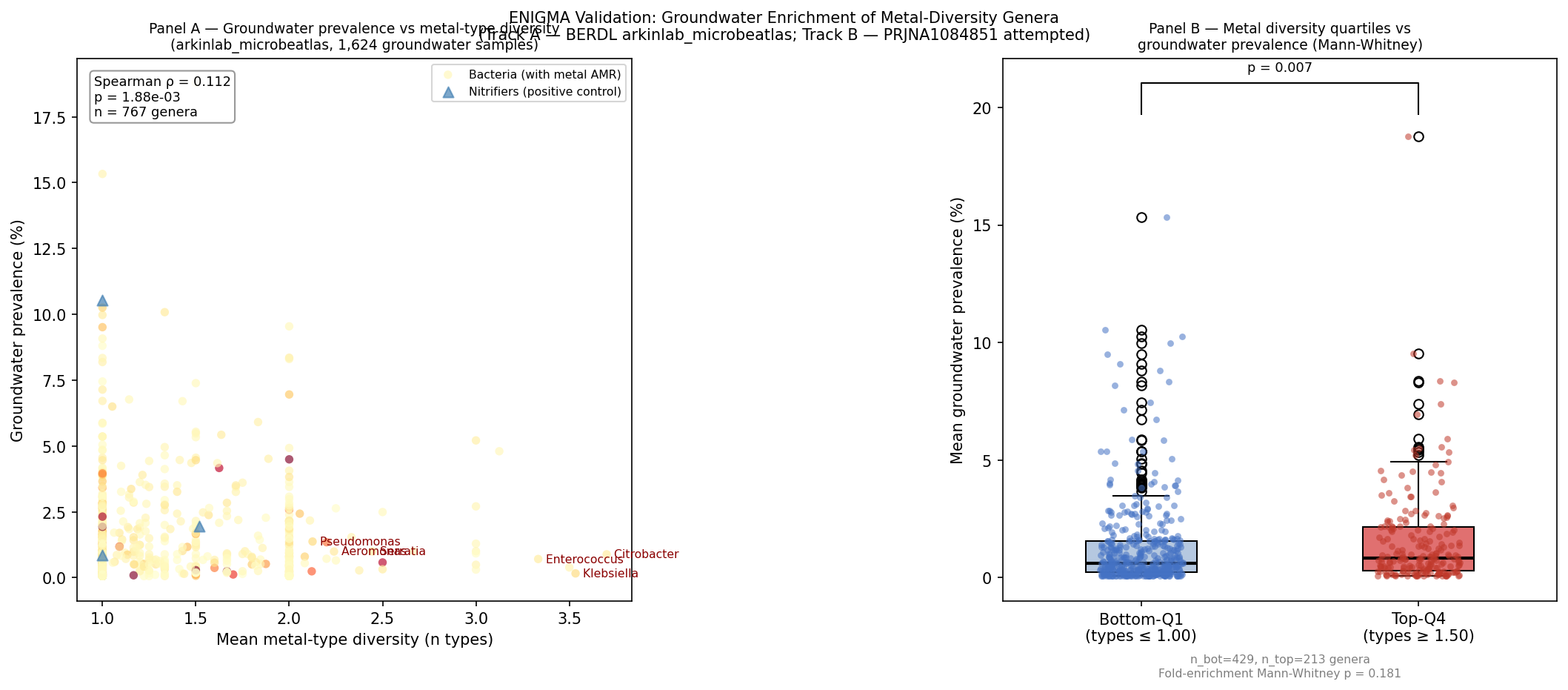
Fig Enigma Validation
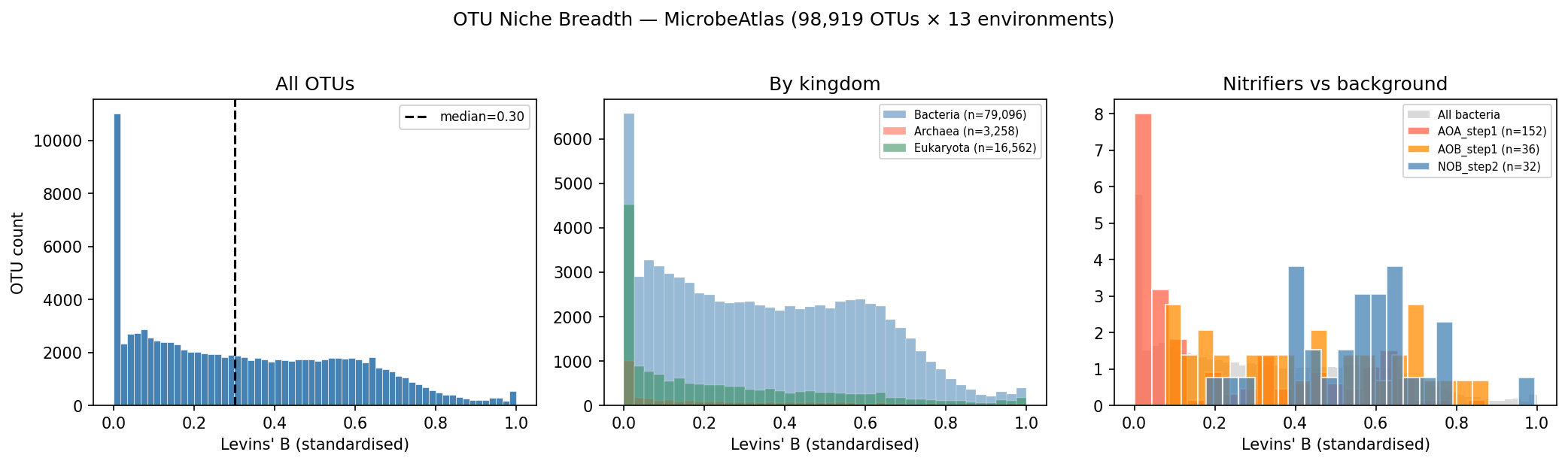
Niche Breadth Distribution
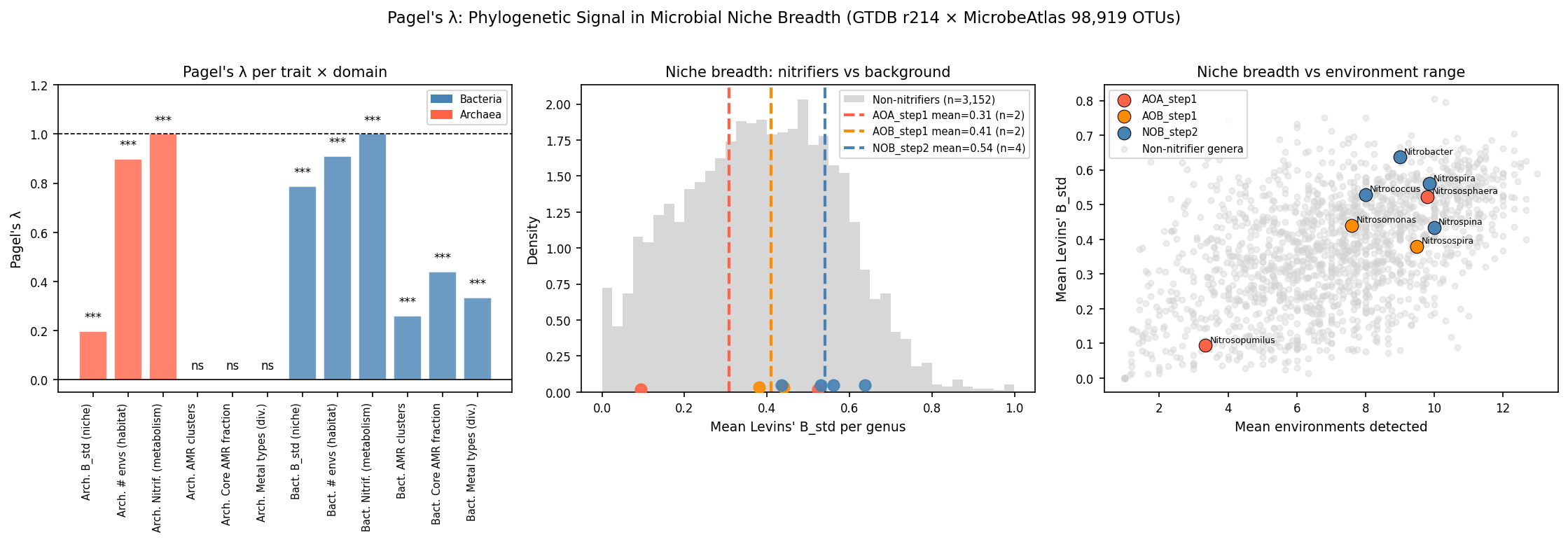
Pagel Lambda Summary
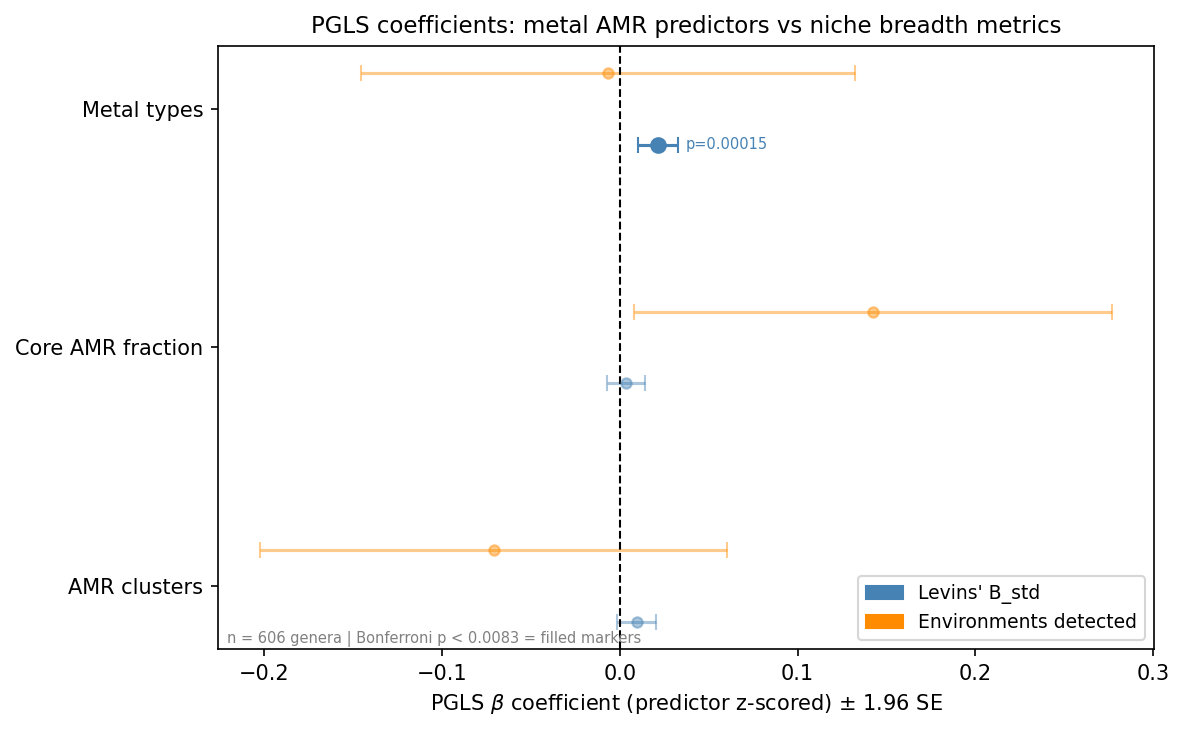
Pgls Forest Plot
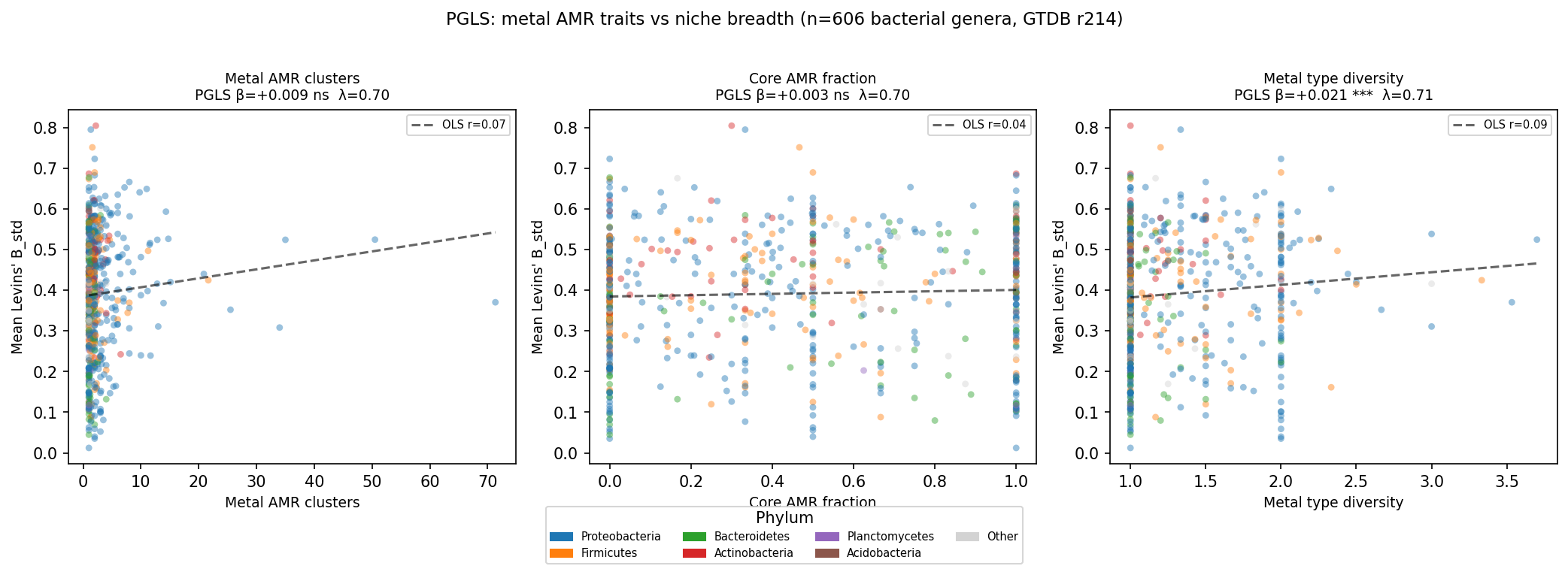
Pgls Scatter
Notebooks
01_metal_amr_species.ipynb
01 Metal Amr Species
View notebook →
02_niche_breadth.ipynb
02 Niche Breadth
View notebook →
03_taxonomy_bridge.ipynb
03 Taxonomy Bridge
View notebook →
04_pagel_lambda.ipynb
04 Pagel Lambda
View notebook →
05_pgls_regression.ipynb
05 Pgls Regression
View notebook →
06_synthesis_figures.ipynb
06 Synthesis Figures
View notebook →
Data Files
| Filename | Size |
|---|---|
all_tests_fdr.csv |
5.4 KB |
enigma_cwm_per_sample.csv |
7.3 KB |
enigma_full_metadata.csv |
19.4 KB |
enigma_geochemistry.csv |
14.9 KB |
enigma_otu_table.csv |
6607.5 KB |
enigma_otu_taxonomy.csv |
1883.7 KB |
enigma_sample_metadata.csv |
4.1 KB |
genus_genome_size.csv |
13.5 KB |
groundwater_enrichment.csv |
61.4 KB |
pgls_3covariate_result.csv |
0.6 KB |
pgls_genome_size_result.csv |
0.4 KB |
pgls_prevalence_threshold_result.csv |
0.2 KB |
pgls_rarefied_summary.csv |
0.3 KB |
pgls_robustness_results.csv |
1.4 KB |
pgls_sensitivity_results.csv |
3.0 KB |
pgls_subset_filtered_otus.csv |
163.8 KB |
prjna1084851_manifest.csv |
5.4 KB |