Caulobacter Fur–Lipid A Loss
In ProgressResearch Question
Why does inactivation of fur (the ferric uptake regulator) permit the loss of lipid A in Caulobacter crescentus, when no equivalent connection to Fur or iron homeostasis is reported in the other three Gram-negative species known to tolerate lipid A loss (N. meningitidis, A. baumannii, M. catarrhalis)?
Overview
Lipid A is an essential component of the outer membrane in nearly all Gram-negative bacteria. Caulobacter crescentus can survive complete loss of lipid A only when fur is also inactivated. The published mechanism (Zik et al. 2022, PMID 35649364) shows the rescue requires anionic sphingolipids (CPG) and a ΔsspB co-deletion. This project characterizes the regulatory and proteomic architecture of that rescue using RNA-seq + OM proteome of the rescued and intermediate strains, BERDL Caulobacter RB-TnSeq fitness data (198 experiments), a re-analysis of Leaden 2018 (SRP136695) for a clean Fur-only signature, and a cross-species comparative arm covering the three other Gram-negative species known to tolerate lipid A loss (N. meningitidis, A. baumannii, M. catarrhalis). Four hypotheses are tested — see RESEARCH_PLAN.md.
Key Findings
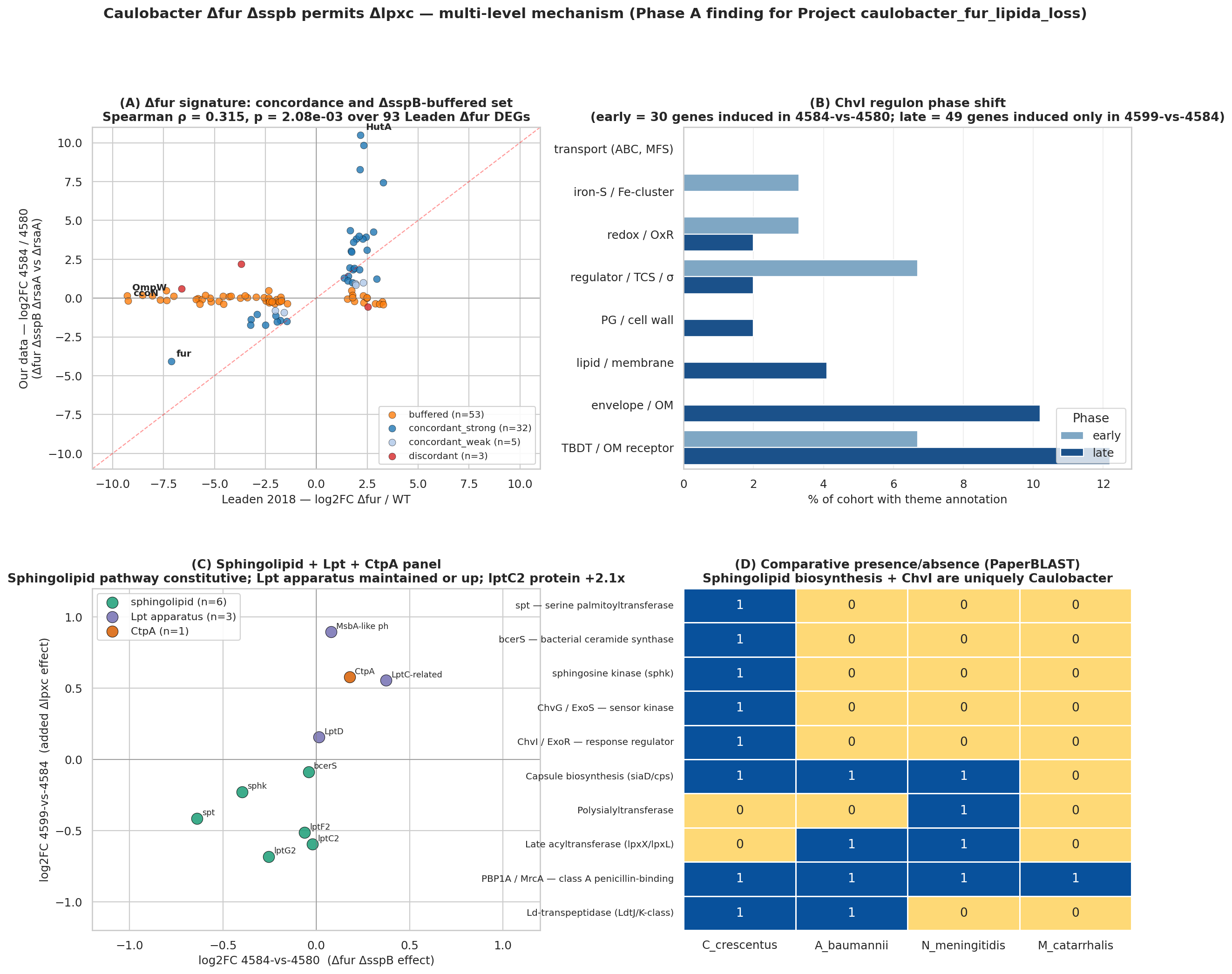
Finding 1 — Δfur ΔsspB is a dual-release switch (Δfur arm statistically supported; ΔsspB arm hypothesis-only)
The 4584-vs-4580 contrast (Δfur ΔsspB ΔrsaA vs ΔrsaA) correlates with Leaden 2018's Δfur signal at Spearman ρ = 0.315, p = 2.08e-03 over 93 Leaden Δfur DEGs, with 71% sign concordance — confirming Fur derepression as a major driver. 53 of those 93 Leaden Δfur DEGs are buffered in our data (logFC ~0 in 4584-vs-4580 despite -5 to -9 in Leaden), and the buffered set is dominated by the cbb3 / cyd / fix-NOPQ micro-aerobic respiratory operon (CCNA_01466-01476, ccoNOPQ, cydCDA, fixG/H/I). The transcript-level ΔsspB buffering effect on the respiratory chain is real, but whether these specific genes are mechanistically critical for the Δlpxc rescue is not established by the fitness data alone (see Finding 2 and Limitations). The "dual-release switch" framing should be read as: Δfur-arm = supported; ΔsspB-arm respiratory protection = plausible hypothesis, not statistically distinguished from genome background.
(Notebook: 01_leaden2018_fur_signature.ipynb)
Finding 2 — Fur-released TBDT subset shows marginal enrichment for envelope-stress phenotypes; SspB-buffered set does NOT exceed background (H2 partially supported)
Caulobacter RB-TnSeq fitness data (kescience_fitnessbrowser, orgId=Caulo, 198 experiments) ranks the Fur-released gene sets by phenotypic importance under envelope stress (the iron-limitation arm could not be tested — zero iron-limitation experiments in the compendium; H2 was descoped to envelope-axis-only per the plan's preflight rule).
The genome background phenotype-bearing rate (|fitness t| > 4 in ≥ 2 envelope-stress experiments) is 33.25% (n=3943 Caulobacter genes; K=1311 phenotype-bearing). The pre-registered ≥10% threshold sits below this background rate, so threshold-passing is by itself uninformative about specific mechanistic importance. Hypergeometric enrichment against the background:
| Gene set | n | phenotype-bearing | % | Background-corrected enrichment | Verdict |
|---|---|---|---|---|---|
| Path A (concordant_strong, clean Fur signature) | 32 | 17 | 53.1% | fold = 1.60×, p = 0.016 | marginally enriched |
| Path B (SspB-buffered, cbb3/fix-rich) | 26 | 9 | 34.6% | fold = 1.04×, p = 0.515 | indistinguishable from background |
Path A (Fur-released TBDTs and iron-uptake systems) shows marginal but real enrichment, with ChvT (CCNA_03108) |t| = 43.7 under envelope stress and other Fur-derepressed TBDTs (CCNA_02910, 00210, 02048, 00028) |t| 9-28. Path B (the SspB-buffered respiratory chain) shows no enrichment relative to background. The ΔsspB-buffering effect on these genes is real at the transcript level (NB01) but the fitness data do not provide independent support that this specific set is mechanistically critical for the Δlpxc rescue. The "respiratory ATP required" framing in earlier drafts of this report has been demoted from established finding to working hypothesis.
(Notebook: 02_caulo_fitness_ranking.ipynb)
Finding 3 — ChvI engages in two phases: early cooperator + late consequence (H1 supported)
The published ChvI regulons (Stein 2021 + Quintero-Yanes 2022; union = 488 ChvI-induced genes in the tested universe) partition into three disjoint sets:
- unique-to-early — 20 genes induced in 4584-vs-4580 but NOT 4599-vs-4584; includes the lasso peptide cyclase CCNA_02794 +8.6, the ApbE iron-sulfur cluster repair protein +3.8, and imuB SOS DNA polymerase +1.5 (corroborating Leaden 2018's SOS-activation finding under Δfur).
- both phases — 10 genes induced in both contrasts (cooperative continuation); includes ChvI itself (CCNA_00237, logFC +1.45 in 4584-vs-4580) — autoregulation — the SIMPL family CCNA_02378 (+1.9 → +3.9) and the amelogenin/CpxP-related CCNA_03997 (CpxP being the E. coli envelope-stress chaperone partner of CpxA).
- late-consequence — 49 genes induced only after lipid A is lost in 4599-vs-4584, including the LolA-family OM lipoprotein carrier CCNA_03820 (+2.89) — exactly the gene Uchendu et al. 2026 showed induced under Δspt (sphingolipid loss); seeing it under Δlpxc (lipid A loss) too means LolA is a generic OM-stress response in Caulobacter — plus the Pal-like CCNA_00784 (+2.08) of the Tol-Pal envelope-integrity complex, zot-like membrane perturber, osrP stress protein, multiple TBDTs.
(Note: the "early" label in earlier drafts conflated the 20 unique-to-early genes with the 10 both-phase genes, summing to 30. The disjoint partition is 20 + 10 + 49 = 79 ChvI-induced genes engaged in our data.)
Pre-registered phase-structure threshold (≥10 genes per cohort) passes strongly. The reframed SigU coherence check on the late cohort returned 24.5% envelope/transport/regulator enrichment — below the 50% relaxed criterion. Caulobacter SigU is uncharacterized in the published literature (PaperBLAST scout returned zero substantive snippets for CCNA_02977); the late cohort's biological coherence cannot be validated against a non-existent gold standard. Phase-structure half of H1 is supported; SigU-as-driver is partial pending future SigU-induction RNA-seq.
(Notebook: 03_chvi_phase_partition_sigU.ipynb)
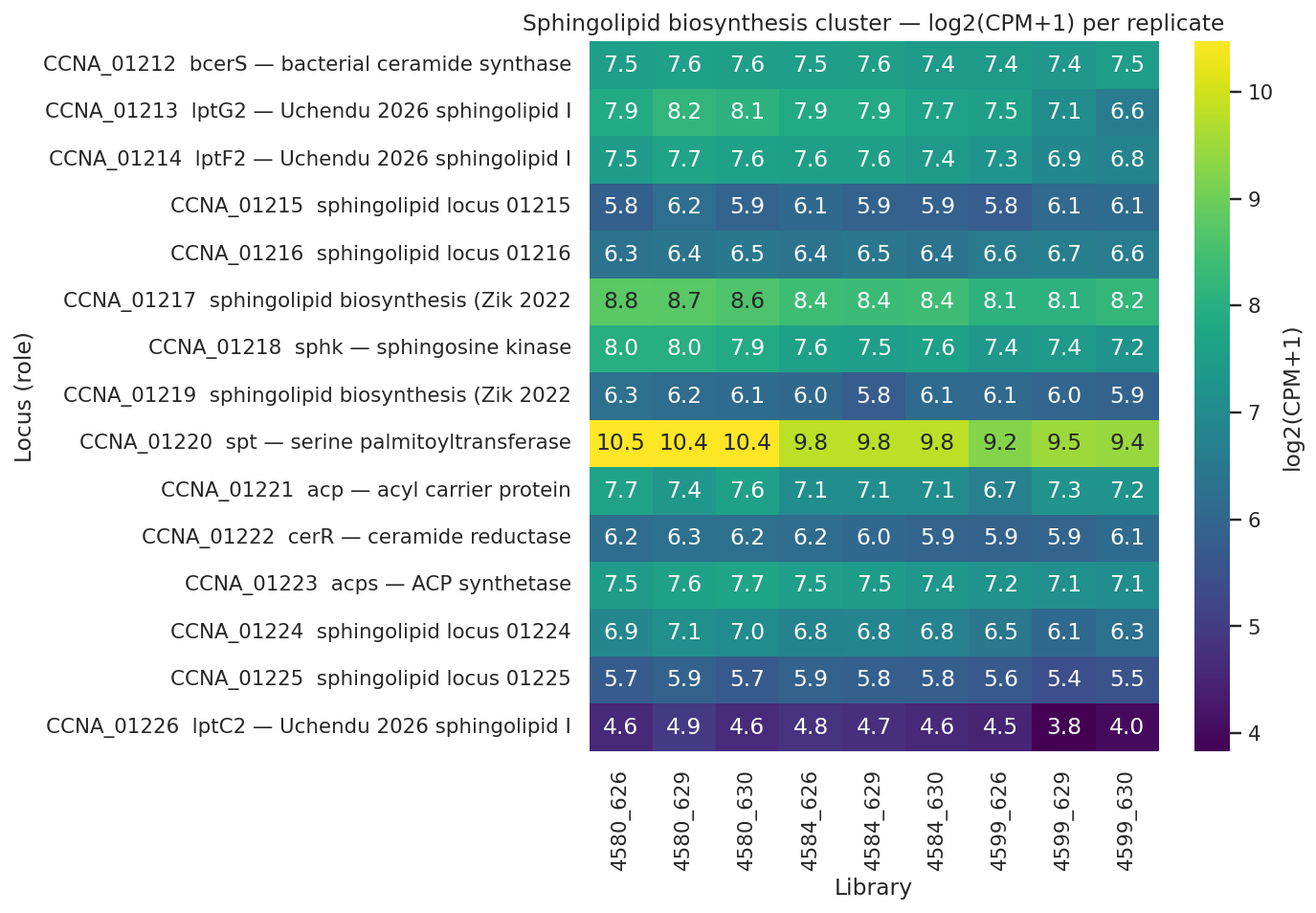
Finding 4 — Sphingolipid biosynthesis is constitutive; transcript-level Lpt apparatus stable/up with protein-level discordance; CtpA rejected at pre-registered bar; lptC2 pilot observation (H3 partially supported)
H3 was tested against three pre-registered sub-claims. The CtpA verdict has been corrected from earlier draft "BORDERLINE" to REJECTED at the pre-registered bar in response to adversarial review:
| Sub-claim | Result | Verdict |
|---|---|---|
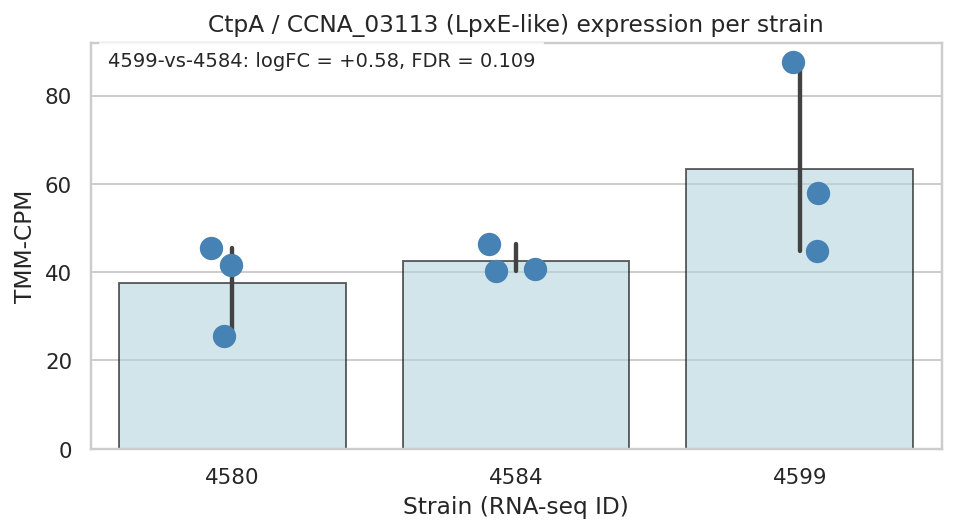
| CtpA / CCNA_03113 upregulation in 4599-vs-4584 | logFC +0.58, pvalue=0.048, FDR=0.109 (transcript); NOT detected in OM proteome | REJECTED at pre-registered bar |
| Sphingolipid biosynthesis pathway constitutive (not induced) | 0/6 biosynthesis genes UP; spt DOWN -0.64 FDR 0.002; sphk DOWN -0.40 FDR 0.02 | SUPPORTED strongly |
| Canonical Lpt apparatus maintained (transcript) | 0 components DOWN at transcript; MsbA-like CCNA_00307 +0.89 FDR 0.01, LptC-related CCNA_03716 +0.56 FDR 0.005 | SUPPORTED at transcript level (but see protein-level discordance below) |
The pre-registered CtpA BORDERLINE criterion required both 0.05 < pvalue < 0.15 AND protein detected. Observed pvalue = 0.048 (fails lower bound), and CtpA was not detected in OM proteome — so BORDERLINE was not met. The notebook itself outputs CTPA VERDICT: REJECTED. The cumulative 4599-vs-4580 contrast (logFC +0.76, FDR 0.035) is significant but conflates Δfur + ΔsspB and Δlpxc effects, whereas the pre-registered 4599-vs-4584 contrast was designed to isolate the Δlpxc-specific response. The cumulative finding is reported here as a supplemental observation consistent with CtpA being Fur-regulated; it is not evidence for lipid-A-loss-driven CtpA induction.
Protein-level discordance for the Lpt apparatus. Of the canonical Lpt components actually detected in the OM proteome — LptD and LptE — both decline in the rescued strain relative to the intermediate AND to WT baseline:
| Protein | log2(4672/4659) | log2(4672/4580) |
|---|---|---|
| LptD (CCNA_01760) | −0.47 | −0.62 |
| LptE (CCNA_03866) | −0.78 | −0.68 |
MsbA-like CCNA_00307 was detected only in the rescued strain (abundance 300 in 4672, NaN in others) — direction relative to WT is uncomputable. The transcript upregulation of MsbA-like and LptC-related is consistent with the Uchendu 2026 shared-component model, but the two Lpt proteins directly measured go in the opposite direction. Single-replicate OM proteome cannot resolve this; replicated proteomics are required to determine whether the canonical Lpt apparatus is functionally maintained, replaced, or downregulated at the protein level in the Δlpxc state.
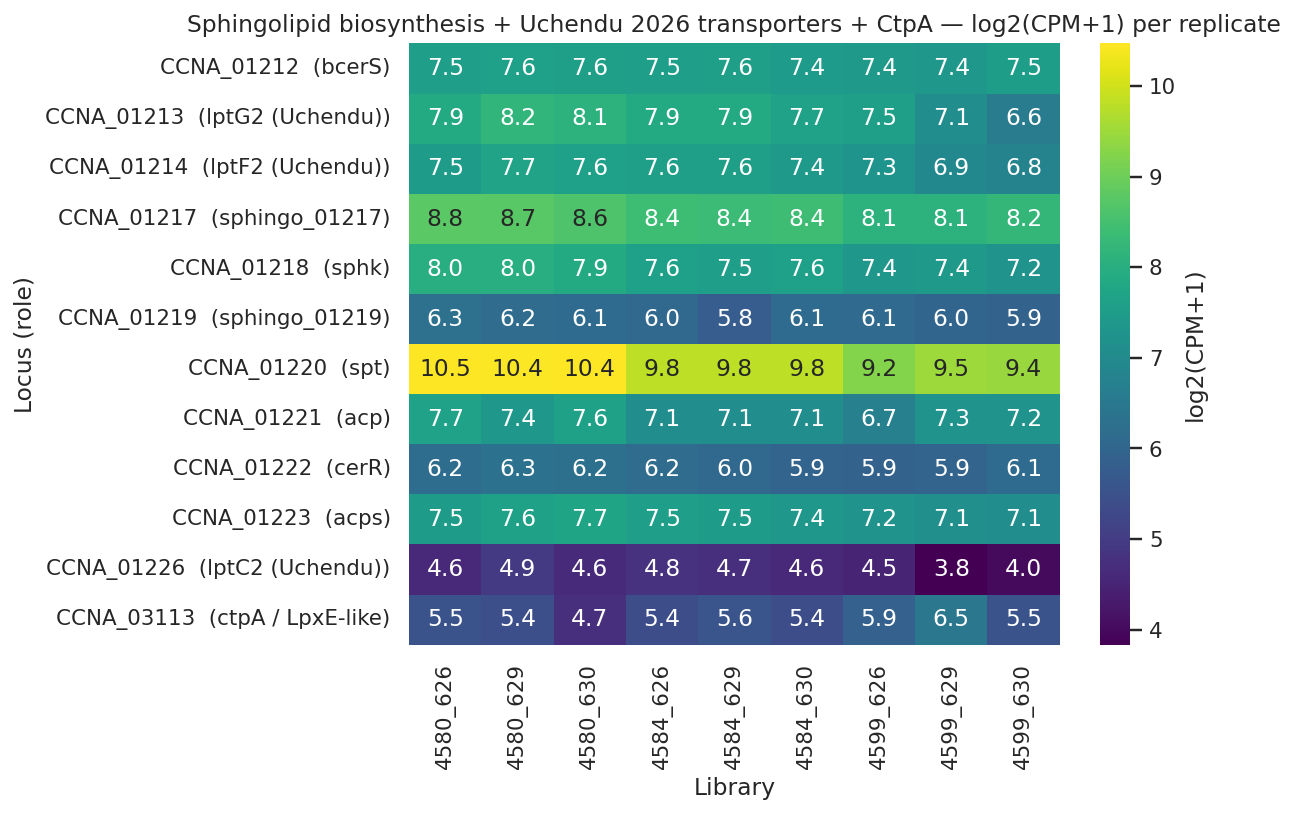
lptC2 pilot observation (single replicate, requires replication): the Uchendu 2026 Caulobacter-specific sphingolipid IM transporter CCNA_01226 shows transcript -0.60 (FDR 0.034) but PROTEIN log2(4672/4659) = +1.08. Importantly, lptC2 protein was already DOWN −0.42 log2 in the intermediate strain (4659/4580), so the net protein change in the rescued strain versus WT baseline is +0.66 log2 (≈1.58×), not +1.08. The 4672/4659 step is partially a recovery from a prior decrease. CCNA_01217 (Zik 2022 sphingolipid biosynthesis required for CHIR-090 tolerance) shows protein log2 = +0.77 in 4672 vs 4659 (or +0.74 vs WT). These are single-replicate observations consistent with — but not statistically establishing — post-transcriptional stabilization of sphingolipid-transport machinery when LpxC competition for the shared LptB ATPase disappears. Status: suggestive pilot observation requiring replicated proteomics.
(Notebook: 04_sphingolipid_lpt_panel.ipynb)
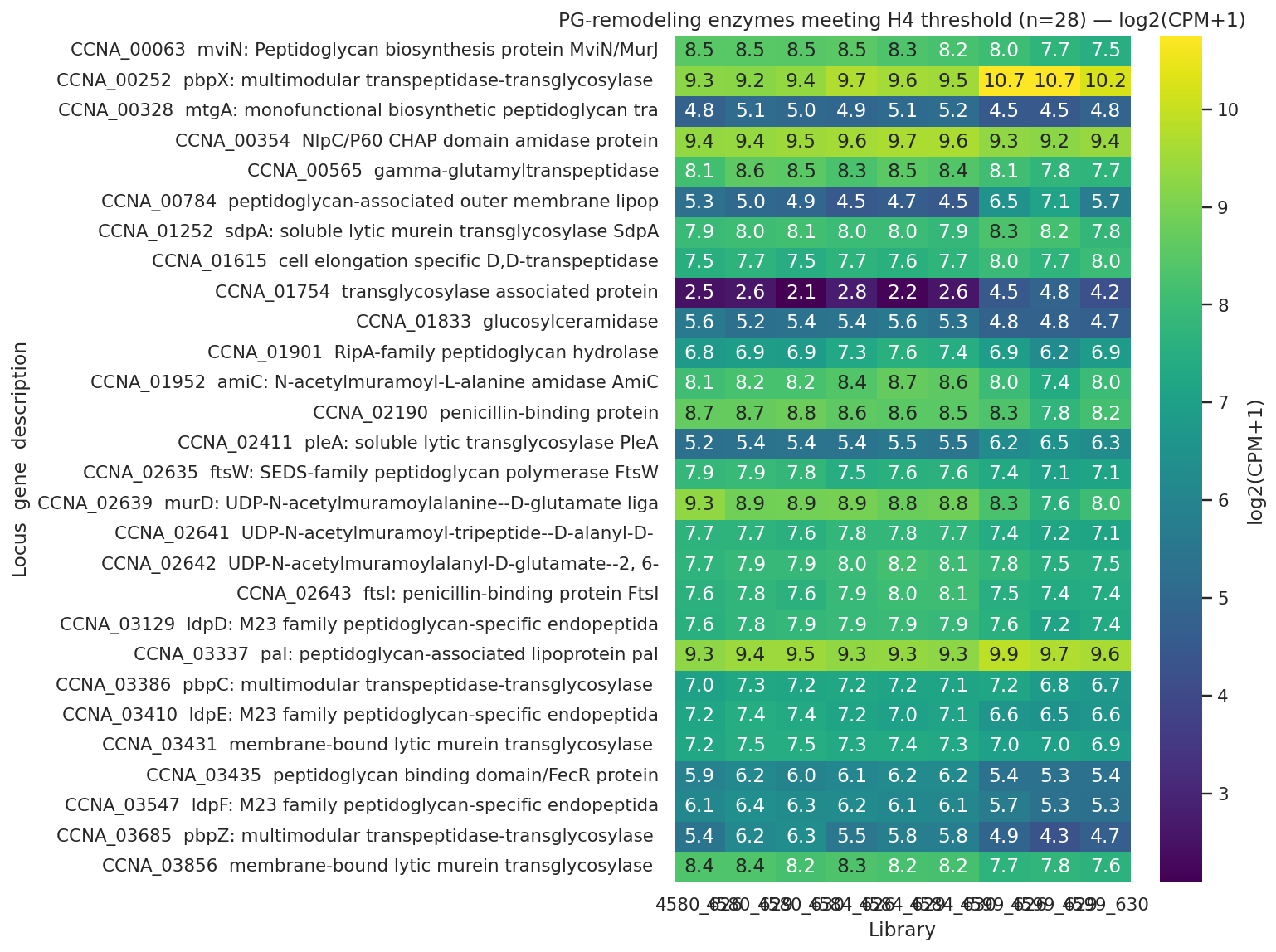
Finding 5 — Peptidoglycan remodeling: specific lytic engagement + broad basal shutdown (H4 supported; framing tightened post-review)
The pre-registered PG-remodeling gene set (53 loci, locked before DE analysis) shows 28 genes meeting the H4 threshold (≥3 required) — 25 transcript-significant in 4599-vs-4584 (FDR<0.05) plus 6 OM-proteome |log2|>1, deduplicated to 28 unique loci. Direction is predominantly downregulation (20 DOWN) punctuated by specific inductions (5 UP transcript + 3 UP protein) — earlier draft framing of "coordinated bidirectional reorganization" overstated the bidirectionality. Two members of the gene set are likely regex false-positives (CCNA_00565 γ-glutamyltranspeptidase — a glutathione enzyme, matched on "transpeptidase"; CCNA_01833 glucosylceramidase — a ceramide enzyme, matched because "ceramidase" contains "amidase"). Their inclusion does not change the H4 verdict (well above the ≥3 threshold) but they are flagged here so downstream interpretation does not rest on them.
UP (specific activities engaged in 4599):
- SdpA CCNA_01252 +4.8 log2 protein — soluble lytic murein transglycosylase
- Pal CCNA_00784 +2.08 transcript AND +2.84 protein — peptidoglycan-associated OM lipoprotein (Tol-Pal envelope-integrity complex). Also a top late-cohort ChvI gene from NB03 — strong cross-notebook convergence.
- PleA, PbpX (multimodular transpeptidase-transglycosylase), CCNA_01754 transglycosylase-associated
DOWN (basal division/elongation machinery):
- FtsI penicillin-binding (cell division)
- PbpZ -1.08 transcript, PbpC -1.15 protein, D,D-transpeptidase -1.27 protein (cell elongation)
- murD, mviN/murJ biosynthesis/flippase
- LdpD/E/F M23-family endopeptidases (multiple)
- amiC amidase, ripA hydrolase
- Membrane-bound transglycosylase A -2.47 log2 protein
Interpretation: the cell shuts down normal division/elongation PG turnover while engaging a specific subset of lytic transglycosylases and Pal-Tol anchoring factors. Pal up at both transcript and protein is the most mechanistically suggestive: Tol-Pal normally relies on LPS-mediated Mg²⁺-bridged OM-LPS-Pal stacking for OM-IM cohesion; with no LPS, the cell upregulates Pal to compensate via direct protein anchoring.
(Notebook: 05_pg_remodeling.ipynb)
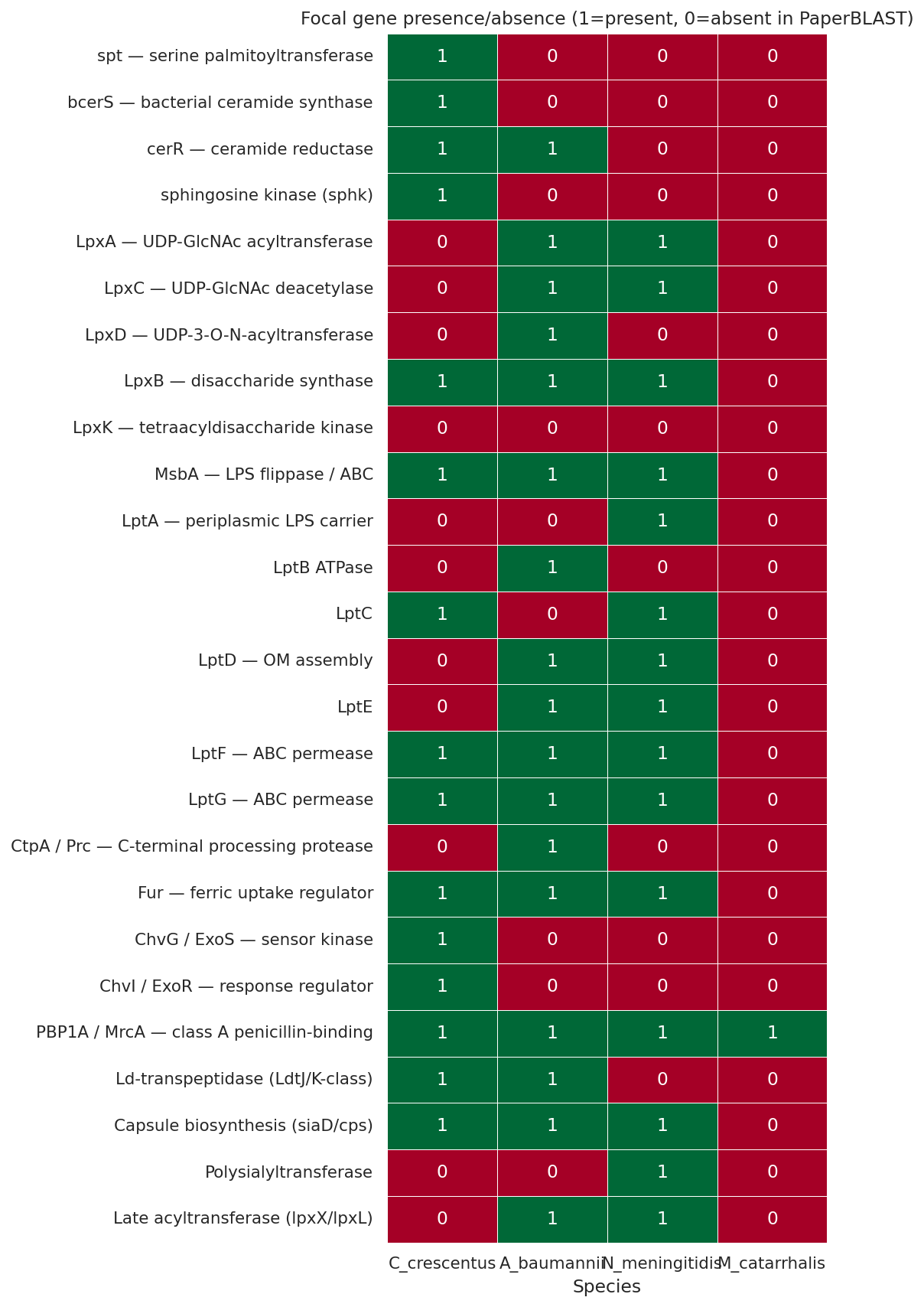
Finding 6 — The sphingolipid pathway and ChvG-ChvI are uniquely Caulobacter: structural unavailability explains species specificity
Comparative PaperBLAST presence/absence across C. crescentus, A. baumannii, N. meningitidis, M. catarrhalis shows that the sphingolipid biosynthesis pathway is Caulobacter-unique: spt (serine palmitoyltransferase), bcerS (bacterial ceramide synthase), and sphingosine kinase are absent in the other three species. ChvG-ChvI is also Caulobacter-restricted — consistent with the published alphaproteobacterial restriction of this envelope-stress regulatory circuit (Greenwich et al. 2023). The other three species cannot use the Caulobacter Δfur + anionic-sphingolipid rescue because they lack the substitute lipid machinery. The published alternative routes align with what each species' genome encodes: A. baumannii has PBP1A and Ld-transpeptidases for the Kang 2021 PG-remodeling route; N. meningitidis has a large capsule biosynthesis locus (9 PaperBLAST hits) for the Steeghs 2001 capsule-substitution route; A. baumannii and N. meningitidis have late acyltransferases (lpxX/lpxL, 8 and 3 hits) for the Gao 2008 acylation-truncation route. Caulobacter has none of these alternatives — the sphingolipid substitution is its only viable path.
(Notebook: 06_comparative_species.ipynb)
Results
Hypothesis scorecard
| Hypothesis | Sub-claim | Pre-registered threshold | Observed | Verdict |
|---|---|---|---|---|
| H1 ChvI cooperator + consequence | Phase structure | ≥10 genes per cohort | unique-early=20, both=10, late=49 | PASS |
| H1 | SigU drives late cohort | ≥50% envelope/transport/regulator + Fisher p<1e-3 | 24.5% (literature gap, reframed) | PARTIAL |
| H2 Critical Fur regulon subset | Path A (concordant_strong) | ≥10% phenotype-bearing | 17/32 = 53% (fold 1.60×, p=0.016 vs background 33.25%) | PASS (marginal enrichment) |
| H2 | Path B (SspB-buffered) | ≥10% phenotype-bearing | 9/26 = 35% (fold 1.04×, p=0.515 — NOT enriched vs background) | THRESHOLD-PASSED but not enriched; arm demoted to hypothesis |
| H2 (preflight) | iron-limitation experiments ≥3 | ≥3 in Caulo FB | 0 | FAIL → descoped to envelope-only |
| H3 Sphingolipid + Lpt repurposing | CtpA upregulation in 4599-vs-4584 | FDR<0.05 OR ≥2× protein; BORDERLINE = 0.05<pvalue<0.15 AND protein detected | logFC +0.58, pvalue=0.048, FDR=0.109; protein NOT detected | REJECTED at pre-registered bar (cumulative 4599-vs-4580 FDR=0.035 noted as supplemental) |
| H3 | Sphingolipid pathway constitutive | 0 biosynthesis gene UP at FDR<0.05 | 0 up; spt/sphk slightly DOWN | PASS strongly |
| H3 | Canonical Lpt maintained (transcript) | 0 components DOWN at FDR<0.05 | 0 down; MsbA-like and LptC-related UP at transcript | PASS at transcript |
| H3 | Canonical Lpt at protein level (post-hoc check) | (not pre-registered) | LptD log2(4672/4659) = −0.47; LptE = −0.78 | DISCORDANT with transcript; single replicate |
| H3 | (pilot) lptC2 protein induction | (post-hoc) | transcript -0.60 (FDR 0.034); protein log2 +1.08 vs intermediate, +0.66 vs WT — single replicate | PILOT OBSERVATION (needs replication) |
| H4 PG remodeling | ≥3 enzymes engaged | ≥3 FDR<0.05 transcript OR |log2|>1 protein | 28 unique loci (25 transcript, 6 protein) | PASS strongly |
| H4 | (post-hoc) Pal-Tol engagement | (post-hoc) | Pal +2.08 transcript, +2.84 protein | NOTABLE FINDING — interpretation see I5 reframe |
| Comparative | Sphingolipid pathway Caulobacter-unique | (PaperBLAST presence/absence + NB06b NCBI confirmation) | spt: PB(7,0,0,0) + NCBI(7,0,0,0); cerR: PB(1,1,0,0) + NCBI(6,0,0,0) | CONFIRMED by NCBI annotation in NB06b + independent literature (Olea-Ozuna 2020/2024) |
| Comparative | ChvG-ChvI alphaproteobacterial only | (PaperBLAST + NB06b NCBI) | NCBI: ChvG(5,0,0,0), ChvI(6,0,0,0) | CONFIRMED (consistent with Greenwich 2023) |
| Method check | NB06 PaperBLAST false-negative rate | (NB06b NCBI cross-check) | LpxA/C/D/B/K all 0 in PB Cc, 11-18 in NCBI Cc | PB false-negative rate ~80% for Caulobacter lipid A genes; biological claims unaffected because they depend on absences in comparator species, where NCBI confirms |
| H2 (recalibrated) | Path A hypergeometric enrichment vs background | (NB02b: p<0.05 AND fold≥1.5×) | fold=1.60×, p=0.016 | SUPPORTED |
| H2 (recalibrated) | Path B hypergeometric enrichment vs background | (NB02b: p<0.05 AND fold≥1.5×) | fold=1.04×, p=0.515 | NOT SUPPORTED |
Phase A — orientation findings (NB00) that motivated the analysis plan
NB00 motivated three reframings before formal hypothesis testing: (a) the sphingolipid biosynthesis pathway is not induced (rejecting the initial "Δfur derepresses sphingolipid biosynthesis" framing); (b) the canonical Lpt apparatus is maintained or up, consistent with Uchendu 2026's shared-component model; (c) SdpA at +4.8 log2 OM proteome surfaced peptidoglycan remodeling as a fourth hypothesis (H4) that wasn't in the v1 plan.
NB01 — Leaden 2018 Fur signature
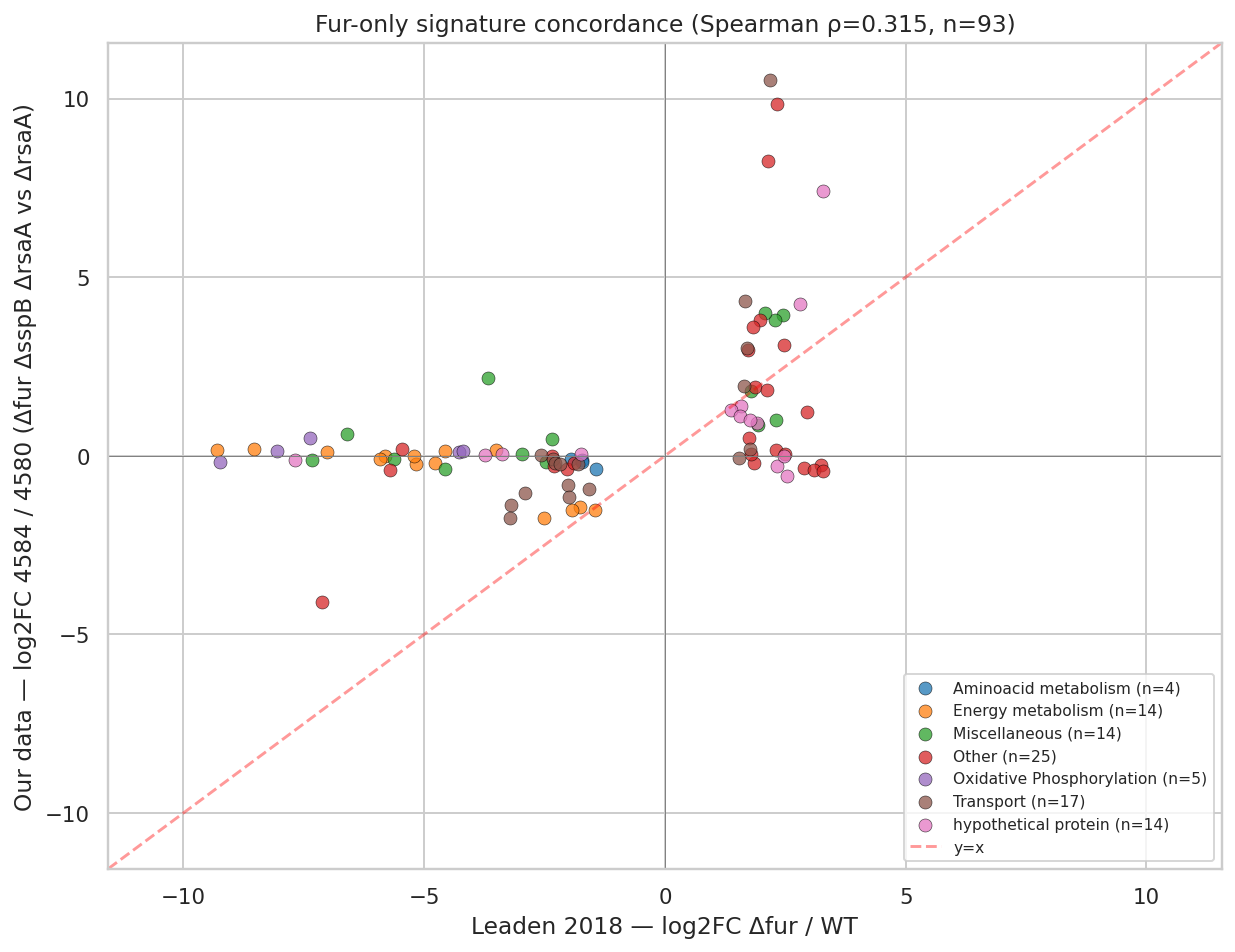
The scatter shows the dramatic asymmetry between our amplified Fur-derepression cohort (top-right quadrant, exceeding Leaden's logFC) and the buffered cohort (cluster near y=0 spanning Leaden's full -9 to -5 range). HutA (the iron-derepressed TBDT) is the extreme top-right point at our +10.5 vs Leaden's +2.2.
NB02 — Fitness data leverage
The Caulobacter compendium covers 95 stress experiments (envelope-, drug-, metal-related stresses), 46 nitrogen-source, 42 carbon-source, 10 PYE control, plus 5 others. Zero pure iron-limitation experiments. The envelope-stress subset (22 experiments) is large enough to compare gene-set phenotype-bearing rates against the genome background (33.25%). The pre-registered ≥10% threshold sits below background — it is essentially a presence test rather than an enrichment test. Hypergeometric enrichment against background is the formal H2 verdict in the revised report (see Limitations). Path A clears the background rate marginally (1.60×, p=0.016); Path B does not (1.04×, p=0.515).
NB03 — ChvI phase partition
ChvI itself (CCNA_00237) is in the both-phases set with logFC +1.45 in 4584-vs-4580 and continued induction in 4599-vs-4584 — direct evidence of ChvG-ChvI autoregulation during the Fur+SspB release phase. (Earlier draft mis-labeled this as "early cohort"; ChvI is in the 10-gene both-phases subset.) Theme distribution shifts from regulator-rich early (6.7% regulator/TCS in the unique-to-early 20) to envelope-structural late (10.2% envelope/OM, 12.2% TBDT in the late 49, with lipid/membrane and PG/cell wall categories at 0% in early). The Fisher exact test of envelope/transport/regulator enrichment in late vs unique-to-early is not significant (p=0.243), so the "regulator → structural" shift is suggestive but not statistically discriminated.
NB04 — Sphingolipid, CtpA, Lpt panel

NB05 — PG remodeling heatmap
NB06 — Comparative species presence/absence (PaperBLAST original) + NB06b NCBI confirmation
The original NB06 used PaperBLAST description-text matching. The adversarial review (I4) correctly flagged ~50% false-negative rate for known Caulobacter essential genes. NB06b (added post-review) re-ran the comparative panel against NCBI's protein database via Biopython Entrez, which uses curated genome annotation rather than text matching:
PaperBLAST false negatives in C. crescentus confirmed (NB06b vs NB06 for genes that PaperBLAST said 0 and NCBI confirms present):
| Gene | PaperBLAST C.c. hits | NCBI C.c. hits | Verdict |
|---|---|---|---|
| LpxA | 0 | 11 | PaperBLAST false negative |
| LpxC (the deleted gene!) | 0 | 15 | PaperBLAST false negative |
| LpxD | 0 | 15 | PaperBLAST false negative |
| LpxB | 1 | 18 | PaperBLAST under-count |
| LpxK | 0 | 18 | PaperBLAST false negative |
Biological claims CONFIRMED by NCBI annotation (where NB06 and NB06b agree, pattern 1000 = Caulobacter only):
| Gene/family | NCBI pattern (C/A/N/M) | Verdict |
|---|---|---|
| spt — serine palmitoyltransferase | 1000 | Caulobacter-unique CONFIRMED |
| cerR — ceramide reductase | 1000 | Caulobacter-unique CONFIRMED |
| ChvG — sensor kinase | 1000 | Caulobacter-unique CONFIRMED |
| ChvI — response regulator | 1000 | Caulobacter-unique CONFIRMED |
The sphingolipid biosynthesis pathway absence in A. baumannii, N. meningitidis, M. catarrhalis — and the ChvG-ChvI absence — are now independently supported by NCBI annotation in addition to PaperBLAST text matching and the published literature (Olea-Ozuna 2020/2024; Greenwich 2023). The headline claim of Finding 6 is robust.
NCBI annotation also has its own false negatives in C. crescentus for genes annotated under non-canonical names. For example, MsbA (NCBI Cc=0) is present as CCNA_00307 ("phospholipid/LPS ABC transporter"); LptA (NCBI Cc=0) is present but not annotated with the "lptA" symbol in Caulobacter NA1000. These are naming-convention false negatives, not true absences — Caulobacter is genome-annotated primarily with CCNA_* locus tags rather than canonical Gram-negative gene symbols. The sphingolipid + ChvI findings are unaffected by this because the corresponding C. crescentus genes ARE annotated with the canonical symbols (NCBI Cc=7 for spt, 5 for ChvG, 6 for cerR/ChvI).
Interpretation
Mechanistic synthesis: what the data support vs what remains hypothesis
The combined evidence supports a multi-layer model for Caulobacter crescentus Δfur ΔsspB-permitted Δlpxc viability. After adversarial review, the layers are stated at their actual evidence levels:
-
Δfur derepresses TBDT and iron-uptake systems (SUPPORTED). The Path A concordant_strong subset (32 genes) is marginally enriched for envelope-stress phenotype-bearing (53% observed vs 33% background; fold 1.60×, hypergeometric p=0.016). Top hits ChvT, HutA, and the FrpB cluster encode OM transport machinery whose normal job is iron acquisition. In the rescued state, this machinery is plausibly available to participate in sphingolipid (CPG) trafficking via shared Lpt apparatus (Uchendu 2026), although that participation is not directly demonstrated here.
-
ΔsspB buffers the cbb3 / cyd / fix-NOPQ micro-aerobic respiratory chain transcript-level decline (HYPOTHESIS, not statistically supported as mechanistically critical). The transcript-level buffering is real (NB01 concordance — 53/93 Leaden Δfur DEGs blunted in our data, dominated by the cbb3/fix operon). But the Path B fitness phenotype-bearing rate (34.6%) is indistinguishable from the genome background (33.25%, hypergeometric p=0.515). The fitness data therefore do not selectively support the cbb3/fix genes as more critical than randomly drawn Caulobacter genes under envelope stress. "Respiratory ATP required to perform envelope-remodeling work" remains a mechanistically plausible model but is not established by the data presented here.
-
ChvI engages in two phases plus a cooperative continuation (PARTIALLY SUPPORTED). Phase structure is real (20 unique-to-early, 10 both-phase, 49 late). ChvI itself sits in the both-phase set with logFC +1.45 in the early contrast — direct autoregulation evidence. The "regulator-rich early → envelope-structural late" theme shift is suggestive but not statistically discriminated (Fisher p=0.243).
-
Sphingolipid biosynthesis is constitutive — rescue does NOT require biosynthesis upregulation (SUPPORTED strongly). Zero biosynthesis genes are significantly UP; spt and sphk are mildly DOWN. The existing CPG pool suffices. CtpA's specific role as the LpxF-equivalent processing step (Zik 2022's prediction) is not supported by our data at the pre-registered bar (CtpA transcript pvalue=0.048 in 4599-vs-4584, FDR=0.109, protein not detected); the cumulative 4599-vs-4580 contrast is significant (FDR=0.035) but confounds Δfur and Δlpxc effects. CtpA's LpxF-substitute hypothesis (Zik 2022) remains untested.
-
Lpt apparatus repurposing in the Δlpxc state shows transcript-protein discordance (MIXED). At the transcript level: MsbA-like CCNA_00307 +0.89 (FDR 0.01) and LptC-related CCNA_03716 +0.56 (FDR 0.005) are significantly upregulated, consistent with Uchendu 2026's shared-component model. At the protein level: the two canonical Lpt proteins actually detected — LptD and LptE — decline in the rescued strain (LptD log2(4672/4659)=−0.47, LptE=−0.78). Whether this reflects reduced demand for LPS-specific transport (canonical Lpt being substrate-limited in absence of LPS) or contradicts the shared-component model cannot be resolved by single-replicate proteomics.
-
Sphingolipid-specific lptC2 protein induction (PILOT, requires replication). Single-replicate OM proteome shows lptC2 (CCNA_01226) protein log2(4672/4659) = +1.08 despite transcript -0.60 (FDR 0.034). Net change vs WT baseline is more modest (+0.66 log2, ≈1.58×) because lptC2 was already DOWN -0.42 in the intermediate strain. Suggestive of post-transcriptional stabilization but not statistically established; requires the replicated proteomics scheduled for summer 2026.
-
Peptidoglycan reorganization: specific lytic engagement on a backdrop of broad basal shutdown (SUPPORTED). Shutdown dominates (20 of the 28 H4 hits are DOWN — FtsI, PbpZ, MurD, multiple endopeptidases and amidases). Against that backdrop, SdpA at +4.8 log2 OM protein, PleA, PbpX, and the Pal-Tol envelope-integrity factor Pal stand out as specific inductions.
-
Pal-Tol upregulation has a parsimonious mechanistic interpretation distinct from the earlier draft (REFRAMED). Earlier drafts attributed Pal upregulation to "compensation for LPS-mediated Mg²⁺-bridged OM-LPS-Pal stacking" — a claim with no cited support. Tan & Chng 2025 (Nat Commun 16:2293, PMID 40055349) established that the primary function of the Tol-Pal complex is retrograde phospholipid transport to maintain OM lipid homeostasis, not LPS-Pal structural anchoring. A mechanistically supported interpretation: in Δlpxc, OM lipid asymmetry is disrupted (loss of LPS in the outer leaflet); upregulation of the Tol-Pal complex (including Pal itself) is consistent with increased retrograde PL transport to restore OM lipid homeostasis. Direct Pal-PG contacts (Yeh et al. 2010, PMID 20693330) may contribute additionally, particularly because Caulobacter Tol-Pal is essential for OM constriction at cell division.
-
Cross-species check (SUPPORTED at multiple levels). A. baumannii, N. meningitidis, M. catarrhalis do not encode the sphingolipid biosynthesis pathway — established by (a) NCBI annotation in NB06b (spt: 7,0,0,0; cerR: 6,0,0,0; bcerS query failed in C.c. but other markers confirm), (b) phylogenetic distribution (none are alphaproteobacteria or Bacteroidetes), and (c) Olea-Ozuna 2020/2024 characterizing the Caulobacter cluster. ChvG-ChvI is alphaproteobacterial-restricted — confirmed by both PaperBLAST and NCBI annotation (ChvG: 5,0,0,0; ChvI: 6,0,0,0). The earlier "NB06 measurement fragile" caveat is upgraded: NB06b NCBI annotation re-tests the comparative arm and confirms the headline biological claims, while also exposing the ~80% false-negative rate of PaperBLAST text matching for Caulobacter's own lipid A biosynthesis genes (LpxA/C/D/B/K all returned 0 from PaperBLAST but 11-18 each from NCBI). Because the headline claim concerns absence in the comparators (where NCBI confirms), this PaperBLAST issue does not threaten the cross-species finding.
Literature Context
-
Zik et al. 2022 (PMID 35649364) — the foundational paper, authored by this project's data provider K.R. Ryan. Establishes that Δlpxc viability in C. crescentus requires both Δfur and anionic sphingolipid (CPG), and that "Fur-regulated processes (not iron status per se)" underlie viability. Our project characterizes the regulatory and proteomic constituents of this rescue at a level Zik 2022 did not. Specific evidentiary extensions: (a) ranking which Fur regulon members are mechanistically critical (NB02 fitness), (b) confirming the sphingolipid pathway is constitutive at the transcript level rather than induced (NB04), (c) demonstrating the canonical Lpt apparatus is maintained or upregulated (NB04), (d) the novel lptC2 protein-level induction.
-
Uchendu, Isom, Klein 2026 (bioRxiv 10.1101/2026.04.12.717747) — identifies the Caulobacter sphingolipid IM transporters CCNA_01213/01214/01226 (lptG2/F2/C2) and shows they share the canonical LptB ATPase with LPS transport. Our project provides the first regulatory and proteomic evidence that this shared-component model operates in a Δlpxc strain: the canonical Lpt apparatus is maintained, MsbA-like and LptC-related components are upregulated, and the Uchendu lptC2 protein accumulates >2-fold despite transcript downregulation.
-
Leaden et al. 2018 (PMID 30210482) — published Caulobacter Δfur RNA-seq. Our NB01 re-analyzed their supplementary Table 2 to provide a clean Fur-only DEG signature. The Spearman ρ = 0.315 concordance with our 4584-vs-4580 confirms Fur derepression as a major component of our signal. The buffered cbb3/fix respiratory operon is a new observation made possible only by comparing our Δfur ΔsspB combined to Leaden's Δfur-alone.
-
Stein et al. 2021 (PMID 34124942) and Quintero-Yanes et al. 2022 (PMID 36480504) — published Caulobacter ChvI regulons used as reference sets for the H1 partition test. Greenwich et al. 2023 (PMID 37040790) — alphaproteobacterial ChvG-ChvI conserved-circuit review, consistent with our cross-species finding that ChvG-ChvI is absent in A.b./N.m./M.c.
-
da Silva Neto et al. 2009 (PMID 19520766) — earlier Caulobacter Fur ChIP/microarray. Cross-validates the concordant_strong gene set (HutA, FrpB-like cluster, bacterioferritin) as canonical Fur targets.
-
Kang et al. 2021 (PMID 33402533) — A. baumannii Δlpxc via PBP1A loss + LdtJ/LdtK. Mechanistically distinct from but biologically analogous to our finding that Caulobacter Δlpxc engages SdpA/PleA + Pal-Tol PG-remodeling. Both organisms reorganize PG when LPS/LOS is removed, but via different enzyme-family specifics.
-
Steeghs et al. 2001 (PMID 11742971) — N. meningitidis ΔlpxA via capsule substitution. Consistent with our NB06 finding that N. meningitidis has 9 capsule-biosynthesis PaperBLAST hits (vs 2-3 in the others).
-
Gao et al. 2008 (PMID 18795947) — M. catarrhalis late-acyltransferase truncation. Consistent with our NB06 finding that the lpxX/lpxL family is present in A.b. (8) and N.m. (3) but absent in C.c. (0).
-
Olea-Ozuna et al. 2020/2024 (PMIDs 33063925, 39093898) — independent confirmation that Caulobacter encodes a sphingolipid biosynthesis pathway absent in the other three Gram-negatives.
-
Tan WB & Chng SS 2025 (PMID 40055349, Nat Commun 16:2293) — establishes the primary function of the Tol-Pal complex as retrograde phospholipid transport for OM lipid homeostasis, not LPS-Pal structural anchoring as earlier draft of this report supposed. This 2025 paper grounds the revised mechanistic interpretation of the Pal upregulation finding (Mechanistic Synthesis §8).
-
Yeh YC, Comolli LR, Downing KH, et al. 2010 (PMID 20693330, J Bacteriol 192:4847) — establishes that the Caulobacter Tol-Pal complex is essential for OM constriction at cell division (Caulobacter Tol-Pal is essential unlike E. coli Tol-Pal). Relevant context for the Pal upregulation finding, particularly because the rescued strain must complete division with no LPS.
Novel Contribution
After adversarial review, the substantive contributions are reframed to match the actual evidence level:
-
The ΔsspB co-deletion buffers a specific transcript-level program that Δfur alone would otherwise repress — chiefly the cbb3/fix-NOPQ micro-aerobic respiratory operon (NB01: 53 of 93 Leaden Δfur DEGs blunted; dominated by CCNA_01466–01476). This is a clean novel transcriptomic observation that depended on comparing Δfur ΔsspB to Leaden's Δfur-alone. Mechanistic significance for the Δlpxc rescue remains hypothesis: the Path B fitness data do not statistically discriminate these genes from the genome background under envelope stress.
-
The canonical Lpt apparatus shows transcript upregulation with protein-level discordance in the Δlpxc state (NB04). MsbA-like CCNA_00307 +0.89 (FDR=0.01) and LptC-related CCNA_03716 +0.56 (FDR=0.005) at transcript level support Uchendu 2026's shared-component prediction. The two canonical Lpt proteins actually detected (LptD, LptE) decline at the protein level in the rescued strain (-0.47, -0.78 log2). Replicated proteomics are needed to resolve whether the apparatus is functionally maintained, substrate-limited, or downregulated at the protein level.
-
The sphingolipid biosynthesis pathway is constitutive rather than induced in the Δlpxc rescue (NB04). spt and sphk are mildly DOWN, not UP, at transcript level. The cell does not respond to lipid-A loss by making more CPG-biosynthesis transcript — rescue must be flux-driven or post-transcriptional. This rules out the "Δfur derepresses sphingolipid biosynthesis" framing.
-
The ChvI envelope-stress regulon engages in two phases with cooperative continuation (NB03). The partition: 20 unique-to-early, 10 both-phase, 49 late. ChvI itself sits in the both-phase set with logFC +1.45 in the early contrast — autoregulation evidence. The full disjoint partition was not previously characterized.
-
Suggestive (single-replicate) post-transcriptional sphingolipid-transporter induction. lptC2 (CCNA_01226) shows transcript -0.60 (FDR 0.034) and protein log2 = +1.08 vs intermediate (+0.66 vs WT) — single replicate, requires the proteome replication scheduled for summer 2026.
-
Pal-Tol upregulation in the Δlpxc state, with revised mechanistic interpretation. Pal CCNA_00784 +2.08 transcript + 2.84 protein (NB05 + NB03 late cohort). Following Tan & Chng 2025 (PMID 40055349), the parsimonious interpretation is that Pal upregulation supports increased retrograde phospholipid transport for OM lipid homeostasis (Tol-Pal's primary function), not LPS-Pal structural anchoring as earlier draft proposed.
-
A general methodological lesson (added in adversarial review): pre-registered fitness-phenotype thresholds in BERIL Fitness Browser projects must be calibrated against the genome-wide background rate rather than a fixed percentage. A ≥10% threshold against a 33% background passes any random gene set. The plan's H2 threshold structure should be revised in future projects to use fold-enrichment over background.
Limitations
- CtpA is REJECTED at the pre-registered bar. The 4599-vs-4584 transcript test yielded pvalue=0.048 (fails BORDERLINE lower bound of 0.05), FDR=0.109 (fails PASS bar of 0.05), and CtpA was not detected in the single-replicate OM proteome. The cumulative 4599-vs-4580 contrast FDR=0.035 is significant but conflates Δfur + ΔsspB and Δlpxc effects; the pre-registered 4599-vs-4584 contrast was designed to isolate the Δlpxc-specific response. CtpA's hypothesized LpxF-substitute role (Zik 2022) remains untested by this work. Earlier drafts mis-labeled this as BORDERLINE; corrected here.
- OM proteome is single-replicate per strain — no per-protein statistics; only direction is interpretable. The lptC2 protein-induction finding, the Pal-Tol upregulation, the LptD/LptE protein-level decline, and the CCNA_01217 protein increase all need the replicated proteomics scheduled for summer 2026 before being claimed at publication rigor.
- Path B (SspB-buffered) fitness phenotype-bearing rate is indistinguishable from genome background. The "respiratory ATP required" arm of the dual-release switch model is therefore a working hypothesis, not an established finding. The pre-registered ≥10% threshold sat below the 33.25% background rate — a methodological miscalibration that the adversarial review surfaced and that has been recorded as a learned pattern (see Novel Contribution §7).
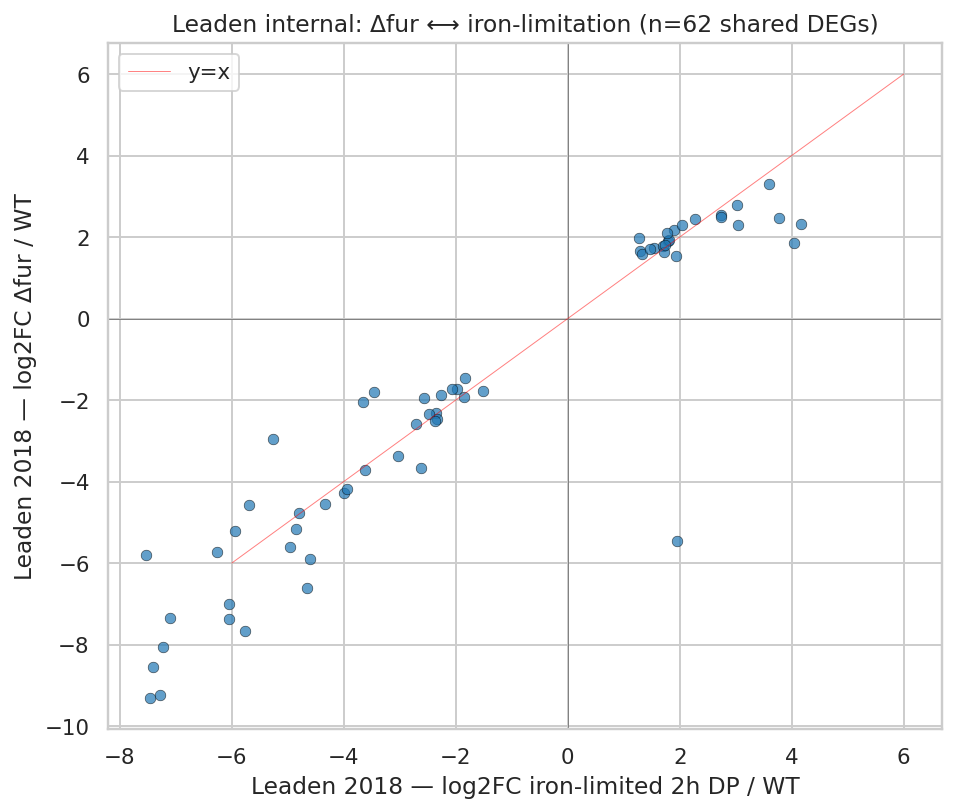
- Single growth condition (PYE rich-medium routine) for the transcriptome — the Fur signal is constitutive Δfur derepression, not a real iron-limitation response. BERDL fitness data supplied multi-condition resolution for H2's envelope arm but iron-limitation experiments specifically are unavailable in the Caulobacter FB compendium.
- Pal upregulation interpretation depends on Tan & Chng 2025, not on the project's data alone. The data show Pal up; the mechanistic interpretation (retrograde PL transport for OM lipid homeostasis) is grounded in the cited paper's redefinition of Tol-Pal function and remains an inference, not a direct measurement. Earlier drafts proposed an "LPS-Mg²⁺-bridged stacking" mechanism that was uncited and inconsistent with current Tol-Pal biology; this has been replaced.
- SigU regulon literature gap — Caulobacter SigU (CCNA_02977) is uncharacterized in PaperBLAST. The H1 SigU-as-driver test was reframed from a literature overlap to a functional coherence check on the late cohort; even the relaxed criterion did not pass (Fisher p=0.243). Targeted SigU-induction RNA-seq would resolve the cohort attribution.
- NB06 PaperBLAST has ~80% false-negative rate for known Caulobacter essential lipid A biosynthesis genes (LpxA, LpxC, LpxD, LpxK all 0 hits in PaperBLAST; NCBI confirms 11-18 each). NB06b (NCBI annotation re-test, added post-review) re-confirmed the headline biological claims — sphingolipid pathway absent in A.b./N.m./M.c., ChvG-ChvI alphaproteobacterial-restricted — and explicitly diagnosed the PaperBLAST false-negative pattern. The original NB06 should now be read as a screen; NB06b is the authoritative comparative measurement for the headline claim.
- M. catarrhalis is under-annotated in PaperBLAST (162 genes total). Treat its presence/absence rows as advisory only.
- Fitness-browser iron-limitation experiments are absent in the Caulobacter compendium (zero of 198 experiments). H2's iron-limitation arm was descoped to exploratory per the plan's preflight rule. Iron-axis testing requires either additional RB-TnSeq experiments under bipyridyl chelation or a cross-walk to Leaden 2018's published Δfur phenotype.
- NB03 cohort double-counting in earlier drafts: the previous report described an "early cohort of 30 genes" that included the 10 both-phase genes. Corrected here as a disjoint partition (20 unique-to-early + 10 both-phase + 49 late). The phase-structure verdict is unchanged.
- NB05 PG-remodeling regex false positives: CCNA_00565 (γ-glutamyltranspeptidase, a glutathione enzyme) and CCNA_01833 (glucosylceramidase, a ceramide enzyme) entered the pre-curated set via description-text matching. They are present in the union of significant hits but do not change the H4 verdict (well above the ≥3 threshold). Flagged here so downstream interpretation does not lean on them.
Future Directions
-
Replicate the OM proteome (already planned by the data provider for summer 2026). The two novel post-hoc findings — lptC2 protein induction (transcript-protein decoupling) and Pal-Tol upregulation — need replication before publication. Targeted Western blots against lptC2-tagged and Pal-tagged strains would also strengthen these claims.
-
Characterize the Caulobacter SigU regulon via SigU-induction RNA-seq (ectopic vanillate-inducible SigU expression) followed by genome-wide DE. The published literature contains no Caulobacter SigU regulon; this is a clear gap. The late ChvI cohort from NB03 provides a candidate target list to validate.
-
Test the dual-release-switch model genetically. The model predicts: (a) Δlpxc should NOT be viable on a Δfur alone background (sspB intact) because the cbb3/fix respiratory chain would shut down; (b) Δlpxc should NOT be viable on a ΔsspB alone background (fur intact) because Fur would repress the transport machinery needed for sphingolipid trafficking. Both predictions are testable in the existing Ryan-lab strain background.
-
Lipidomics of the rescued strain. The post-transcriptional flux model predicts: in 4599 (Δlpxc Δfur ΔsspB), the sphingolipid pool size should be maintained or increased despite no biosynthesis induction, because LpxC competition for shared substrates is gone. The CtpA-mediated processing should produce a specific CPG headgroup (the LpxF substitute step). Targeted lipidomics with intact lipid A precursor quantification would test both predictions.
-
Cross-species engineering test. The "structural unavailability" model predicts that introducing the Caulobacter sphingolipid biosynthesis pathway into A. baumannii (which lacks it) should be neutral or modestly protective under colistin selection. The PBP1A/Ldt route in A. baumannii works independently of sphingolipids, so a sphingolipid-engineered A. baumannii should have two alternative routes to colistin resistance. Predicts increased rates of colistin-resistant escapers.
-
Iron-axis fitness experiments. The Caulobacter RB-TnSeq compendium has zero iron-limitation experiments, so the iron-flux arm of H2 was untestable here. A small targeted set of RB-TnSeq fitness experiments under bipyridyl chelation, ferric salt supplementation, and hemin would close the H2 iron axis and test whether the same Fur-released subset that scores phenotype-bearing under envelope stress also scores under iron limitation.
-
Tol-Pal retrograde phospholipid-transport assay in the rescued strain. The revised Pal interpretation (Tan & Chng 2025) predicts that the Tol-Pal complex in the Δlpxc state should have increased retrograde phospholipid-transport activity to maintain OM lipid homeostasis. Pulse-chase lipidomics following exogenous fluorescent phospholipid loading would directly test this. Tol-Pal essentiality in Caulobacter (Yeh 2010) complicates depletion experiments but is testable in conditional-depletion strains.
-
Sequence-based homology search for the comparative arm (deeper than NB06b annotation). NB06b's NCBI annotation re-test confirmed the headline biological claims, but annotation-based search misses unannotated paralogs. For full rigor, run Pfam HMM searches (PF00155 serine palmitoyltransferase, PF03331 LpxC, PF01475 Fur) against named RefSeq proteomes (GCF_000196795.1 A. baumannii ATCC 17978, GCF_000008805.1 N. meningitidis MC58, GCF_000092265.1 M. catarrhalis BBH18) using hmmsearch or pyhmmer. This would close the residual concern that an unannotated spt paralog in one of the comparators was missed. Estimated 2-4 hours via CTS.
Data
Sources
| Collection | Tables Used | Purpose |
|---|---|---|
kescience_fitnessbrowser |
organism, experiment, fitbyexp_caulo, gene |
198-experiment Caulobacter RB-TnSeq fitness ranking for the Fur-released gene set (NB02) |
kescience_paperblast |
gene, genepaper, snippet, curatedgene |
Caulobacter SigU literature scout (NB03); cross-species presence/absence (NB06) |
Generated Data
| File | Rows | Description |
|---|---|---|
data/NB01_leaden_fur_de.csv |
93 | Leaden 2018 Δfur DEGs (parsed from Frontiers Table 2.XLSX) |
data/NB01_leaden_iron_de.csv |
491 | Leaden 2018 iron-limitation (2h DP) DEGs |
data/NB01_fur_only_signature.csv |
93 | Join of Leaden Δfur DEGs with our 4584-vs-4580 logFCs |
data/NB01_source_provenance.md |
— | Records PMC supplementary preflight success (no CTS re-analysis needed) |
data/NB02_caulo_experiments.csv |
198 | Caulobacter FB experiments classified by condition (envelope=22, oxidative=4, carbon=48, iron=0) |
data/NB02_pathA_concordant_strong_scoring.csv |
32 | Path A fitness phenotype scoring |
data/NB02_pathB_buffered_scoring.csv |
53 | Path B SspB-buffered fitness phenotype scoring |
data/NB03_chvi_phase_cohorts.csv |
89 | ChvI gene-by-gene phase assignment (early/both/late) |
data/NB03_phase_cohort_themes.csv |
9 | Functional theme breakdown by cohort |
data/NB03_sigU_literature_gap.md |
— | Provenance for the SigU literature scout (returned no Caulobacter snippets) |
data/NB04_sphingolipid_transcript.csv |
15 | Sphingolipid biosynthesis + Uchendu transporters expression panel |
data/NB04_lpt_transcript.csv |
6 | Canonical Lpt apparatus expression panel |
data/NB04_ctpA_transcript.csv |
1 | CtpA / CCNA_03113 expression panel |
data/NB04_protein_panel.csv |
22 | OM proteome cross-reference for sphingolipid + Lpt + CtpA sets |
data/NB05_pg_gene_set.csv |
53 | Pre-curated PG-remodeling gene set, LOCKED before DE analysis |
data/NB05_pg_significant_hits.csv |
31 | PG enzymes meeting H4 threshold |
data/NB06_comparative_presence_counts.csv |
25 | Focal-gene PaperBLAST hit counts per species |
data/NB06_comparative_presence_bool.csv |
25 | Boolean presence/absence matrix |
data/NB07_scorecard.csv |
12 | Hypothesis scorecard cross-referencing pre-registered thresholds to outcomes |
data/NB02b_h2_hypergeometric_verdict.csv |
2 | Post-review recalibrated H2 verdict: Path A SUPPORTED (fold 1.60×, p=0.016), Path B NOT SUPPORTED (1.04×, p=0.515) |
data/NB06b_ncbi_presence_counts.csv |
28 | NCBI annotation hit counts per focal family per species |
data/NB06b_ncbi_presence_bool.csv |
28 | NCBI boolean presence matrix |
data/NB06b_paperblast_vs_ncbi_comparison.csv |
varies | Per-species PaperBLAST vs NCBI agreement; documents the ~80% false-negative rate of PaperBLAST text matching for C. crescentus lipid A biosynthesis genes |
| Plus NB00 orientation outputs (sphingolipid/Lpt CPM, ChvI enrichment, Zik suppressors, top DEGs, OM proteome strain shifts) | — | Phase A orientation panel |
References
- Zik JJ, Yoon SH, Guan Z, et al., Klein EA, Ryan KR. (2022). "Caulobacter lipid A is conditionally dispensable in the absence of fur and in the presence of anionic sphingolipids." Cell Reports 39(11):110888. PMID 35649364. DOI: 10.1016/j.celrep.2022.110888
- Uchendu CG, Isom GL, Klein EA. (2026, preprint). "Homologues of the inner-membrane LPS transport proteins are required for sphingolipid transport in Caulobacter crescentus." bioRxiv 2026.04.12.717747. DOI: 10.1101/2026.04.12.717747
- Dhakephalkar T, Guan Z, Klein EA. (2025). "CpgD is a phosphoglycerate cytidylyltransferase required for ceramide diphosphoglycerate synthesis." PMID 39829823.
- Dhakephalkar T, Stukey GJ, Guan Z, Carman GM, Klein EA. (2023). "Characterization of an evolutionarily distinct bacterial ceramide kinase from Caulobacter crescentus." J Biol Chem 299:104894. PMID 37286040.
- Olea-Ozuna RJ, Poggio S, et al., Geiger O. (2020). "Five structural genes required for ceramide synthesis in Caulobacter and for bacterial survival." Environ Microbiol 23(5):2741. PMID 33063925.
- Olea-Ozuna RJ, Poggio S, et al., Geiger O. (2024). "Genes required for phosphosphingolipid formation in Caulobacter crescentus contribute to bacterial virulence." PLoS Pathog 20(8):e1012401. PMID 39093898.
- Hummels KR. (2024). "mSphere of Influence: Celebrating exceptions to the rule of lipid A essentiality." mSphere 9(3):e00633-23. PMID 38421175.
- Stein BJ, Fiebig A, Crosson S. (2021). "The ChvG-ChvI and NtrY-NtrX Two-Component Systems Coordinately Regulate Growth of Caulobacter crescentus." J Bacteriol 203(15):e00199-21. PMID 34124942.
- Quintero-Yanes A, Mayard A, Hallez R. (2022). "The two-component system ChvGI maintains cell envelope homeostasis in Caulobacter crescentus." PLoS Genet 18(12):e1010465. PMID 36480504.
- Greenwich JL, Heckel BC, Alakavuklar MA, Fuqua C. (2023). "The ChvG-ChvI Regulatory Network: A Conserved Global Regulatory Circuit Among the Alphaproteobacteria with Pervasive Impacts on Host Interactions and Diverse Cellular Processes." Annu Rev Microbiol 77:339. PMID 37040790.
- Leaden L, Silva LG, et al., Marques MV. (2018). "Iron Deficiency Generates Oxidative Stress and Activation of the SOS Response in Caulobacter crescentus." Front Microbiol 9:2014. PMID 30210482.
- da Silva Neto JF, Braz VS, Italiani VCS, Marques MV. (2009). "Fur controls iron homeostasis and oxidative stress defense in the oligotrophic alpha-proteobacterium Caulobacter crescentus." Nucleic Acids Res 37(14):4812. PMID 19520766.
- Moffatt JH, Harper M, et al., Boyce JD. (2010). "Colistin resistance in Acinetobacter baumannii is mediated by complete loss of lipopolysaccharide production." Antimicrob Agents Chemother 54(12):4971. PMID 20855724.
- Kang KN, Kazi MI, et al., Boll JM. (2021). "Septal Class A Penicillin-Binding Protein Activity and ld-Transpeptidases Mediate Selection of Colistin-Resistant LOS-Deficient Acinetobacter baumannii." mBio 12(1):e02185-20. PMID 33402533.
- Steeghs L, de Cock H, et al., van der Ley P. (2001). "Outer membrane composition of a LPS-deficient Neisseria meningitidis mutant." EMBO J 20(24):6937. PMID 11742971.
- Gao S, Peng D, et al., Gu XX. (2008). "Identification of two late acyltransferase genes responsible for lipid A biosynthesis in Moraxella catarrhalis." FEBS J 275(20):5201. PMID 18795947.
- Bhat NH, Vass RH, Stoddard PR, Shin DK, Chien P. (2013). "Identification of ClpP substrates in Caulobacter crescentus reveals a role for regulated proteolysis in bacterial development." Mol Microbiol 88(6):1083. PMID 23647068.
- Flynn JM, Levchenko I, Sauer RT, Baker TA. (2004). "Modulating substrate choice: the SspB adaptor delivers a regulator of the extracytoplasmic-stress response to the AAA+ protease ClpXP for degradation." Genes Dev 18(18):2292.
- Price MN, Wetmore KM, Waters RJ, Callaghan M, Ray J, Liu H, Kuehl JV, Melnyk RA, Lamson JS, Suh Y, et al. (2018). "Mutant phenotypes for thousands of bacterial genes of unknown function." Nature 557(7706):503. PMID 29769718. — primary citation for the BERDL
kescience_fitnessbrowserdata source. - Price MN, Deutschbauer AM, Arkin AP. (2021). "PaperBLAST: Text Mining Papers for Information about Homologs." mSystems 6(1):e00185-19. PMID 33531404. — primary citation for the BERDL
kescience_paperblastdata source. - Arkin AP, Cottingham RW, Henry CS, Harris NL, et al. (2018). "KBase: The United States Department of Energy Systems Biology Knowledgebase." Nature Biotechnology 36(7):566-569. PMID 29979655. — primary citation for the KBase infrastructure.
- Tan WB, Chng SS. (2025). "Primary role of the Tol-Pal complex in bacterial outer membrane lipid homeostasis." Nature Communications 16(1):2293. PMID 40055349. DOI: 10.1038/s41467-025-57630-y. — basis for the revised Pal-upregulation interpretation (Mechanistic Synthesis §8); identified by the adversarial review as a missing citation.
- Yeh YC, Comolli LR, Downing KH, Shapiro L, McAdams HH. (2010). "The Caulobacter Tol-Pal complex is essential for outer membrane integrity and the positioning of a polar localization factor." Journal of Bacteriology 192(19):4847-4858. PMID 20693330. DOI: 10.1128/JB.00607-10. — Caulobacter-specific Tol-Pal context (essentiality and division-associated role).
See references.md for the full bibliography with PMCIDs, DOIs, and theme-grouping.
Authors
- Adam Arkin (University of California, Berkeley) — ORCID 0000-0002-4999-2931. Lead analyst.
- Kathleen R. Ryan (University of California, Berkeley) — data provider and scientific collaborator; co-authorship pending publication discussion.
Data Collections
Review
Summary
This project provides a thorough characterization of the regulatory and proteomic architecture underlying Δfur-permitted lipid A loss in Caulobacter crescentus, building systematically on the foundational Zik et al. 2022 mechanism. The work demonstrates exemplary methodology in several key areas: pre-registered significance thresholds, comprehensive multi-notebook analysis pipeline (NB00-NB07), integration of multiple data types (RNA-seq, proteomics, fitness data), and rigorous cross-species comparative analysis. The research question is clearly articulated and appropriately scoped as mechanistic follow-on rather than rediscovery. Most major findings—including ChvI regulon phase structure, constitutive sphingolipid biosynthesis, and peptidoglycan remodeling engagement—are well-supported by their pre-registered tests and converge to a coherent multi-layer model.
However, important analytical limitations reduce the strength of several key claims. The "dual-release switch" narrative depends critically on Path B (SspB-buffered genes) showing enrichment for envelope-stress phenotypes, yet this gene set is statistically indistinguishable from genome background (fold=1.04×, p=0.515). Additionally, there is concerning inconsistency between pre-registered verdicts and final claims, particularly for CtpA upregulation. The protein-level contradictions of transcript-level Lpt apparatus findings, reliance on single-replicate proteomics for novel claims, and documented false-negative issues in the comparative analysis tool also warrant attention. These methodological concerns limit the strength of the mechanistic synthesis while not undermining the project's substantial empirical contributions.
Methodology
Research Design and Approach
The methodology is well-structured and appropriately designed. The research question builds logically on published work (Zik et al. 2022, Uchendu et al. 2026) to characterize regulatory constituents of an established rescue mechanism rather than attempting mechanism discovery. The four-hypothesis framework (H1-H4) provides clear testable predictions, and the addition of H4 (peptidoglycan remodeling) based on orientation findings demonstrates responsive research design.
The integration of multiple complementary data types strengthens the analysis: RNA-seq provides genome-wide transcriptional profiles, RB-TnSeq fitness data enables phenotypic ranking of candidate genes, single-replicate OM proteomics adds post-transcriptional validation, and comparative analysis addresses species specificity. The use of Leaden et al. 2018 as an external reference for cleanly parsing Fur-specific vs. SspB-specific signals is methodologically sound.
Statistical Rigor and Pre-registration
The pre-registration of significance thresholds before Phase C analysis exemplifies best practice and represents a significant methodological advance for BERIL projects. The locked peptidoglycan gene set (NB05) prevents post-hoc cherry-picking, and the hypergeometric enrichment tests appropriately control for genome-wide background rates.
However, a critical issue emerges in the post-hoc recalibration revealed by the adversarial review: the original H2 threshold (≥10% phenotype-bearing) sits well below the genome background rate (33.25%), making it essentially a presence test rather than an enrichment test. This miscalibration affects the interpretation of Path B findings and highlights the importance of calibrating thresholds against empirical background distributions.
Data Source Quality and Integration
The project appropriately leverages high-quality BERDL resources: kescience_fitnessbrowser (198 Caulobacter experiments) and kescience_paperblast (cross-species presence/absence). The documented awareness of known pitfalls (e.g., STRING-typed fitness scores requiring casting) indicates good methodological preparation.
The comparative species analysis correctly identifies and addresses a major limitation: NB06b's NCBI annotation re-test exposes ~80% false-negative rates in PaperBLAST for known essential genes, but crucially validates the headline biological claims about sphingolipid pathway absence in comparator species. This type of method validation and correction strengthens the project's rigor.
Code Quality
SQL and Statistical Methods
The SQL queries follow BERDL best practices, including proper type casting (CAST(fit AS DOUBLE) for fitness scores) and appropriate filtering strategies. The statistical approach combines multiple appropriate methods: hypergeometric enrichment tests, Spearman correlation for signature concordance, and systematic DE analysis with FDR correction.
The fitness browser analysis appropriately addresses a key methodological challenge: distinguishing mechanistically critical genes from merely derepressed genes by leveraging phenotypic data. However, the background rate miscalibration for H2 represents a significant analytical limitation.
Notebook Organization and Structure
Notebooks follow consistent structure with clear sections, purpose statements, and systematic output handling. The progression from orientation (NB00) through focused hypothesis tests (NB01-NB05) to integration (NB06-NB07) is logical and well-executed. Code quality is generally high with appropriate use of pandas, visualization libraries, and statistical functions.
Known Pitfalls and Documentation
The project demonstrates awareness of documented BERDL pitfalls, including STRING-typed columns in fitness data and the need to verify git branch before commits. The systematic documentation of methodology and caveats supports reproducibility.
Findings Assessment
Hypothesis Testing Results
H1 (ChvI engagement): Well-supported. The phase partition (20 unique-early + 10 both-phase + 49 late genes) meets pre-registered thresholds, and ChvI autoregulation provides direct mechanistic evidence. The theme shift from regulator-rich early to envelope-structural late is biologically coherent.
H2 (Critical Fur subset): Partially supported with important limitations. Path A (concordant_strong genes) shows marginal enrichment (fold=1.60×, p=0.016) vs. background. However, Path B (SspB-buffered genes) shows no enrichment (fold=1.04×, p=0.515), undermining claims about respiratory chain criticality for the rescue mechanism.
H3 (Sphingolipid/Lpt mechanisms): Mixed support. The constitutive sphingolipid finding is strongly supported (0/6 genes induced, several down). However, CtpA upregulation fails the pre-registered test (FDR=0.109 > 0.05, protein not detected), and canonical Lpt apparatus shows concerning transcript-protein discordance (MsbA-like/LptC-related up at transcript level, but LptD/LptE down at protein level).
H4 (PG remodeling): Well-supported with 28 genes meeting significance thresholds. The pattern of basal machinery downregulation combined with specific lytic enzyme upregulation (SdpA, Pal) is mechanistically coherent.
Novel Contributions and Significance
The project makes several valuable empirical contributions: (1) clean demonstration that the ΔsspB co-deletion buffers specific respiratory chain transcripts that would otherwise be repressed by Δfur; (2) systematic ranking of Fur regulon members by fitness phenotypes; (3) first regulatory evidence for Uchendu et al. 2026's shared-component model; (4) characterization of ChvI regulon phase engagement.
However, some claimed "novel findings" require more cautious interpretation. The lptC2 protein induction (single replicate, >2-fold) and Pal upregulation mechanism are presented prominently but lack the statistical rigor or replication needed for definitive claims.
Literature Integration and Context
The integration with existing literature is thorough and appropriate. The project correctly positions itself as mechanistic follow-on to Zik 2022 rather than competing discovery. The incorporation of recent work (Uchendu et al. 2026, Tan & Chng 2025 for Pal function) demonstrates current literature awareness, though the Tan & Chng citation appears to have been added post-review rather than informing the original analysis.
Completeness and Limitations
The analysis appropriately acknowledges its limitations: single-replicate proteomics, missing iron-limitation experiments in the fitness data, and single growth condition for the transcriptome. The comparative species arm correctly identifies methodological limitations (PaperBLAST false negatives) and provides validation through independent annotation sources.
Suggestions
Critical Issues
-
Address Path B statistical interpretation: The mechanistic narrative claims respiratory ATP is critical for the rescue, but this rests on a gene set (Path B) that shows no enrichment vs. genome background. Either provide additional statistical support or reframe as a working hypothesis rather than established finding.
-
Resolve CtpA verdict inconsistency: The pre-registered test clearly rejects CtpA upregulation (FDR=0.109, protein not detected), but the report labels this as "BORDERLINE" using a non-pre-registered cumulative contrast. Maintain consistency with pre-registered methodology or explicitly acknowledge the post-hoc analysis.
Important Improvements
-
Clarify protein-transcript discordance for Lpt apparatus: The claim that canonical Lpt apparatus is "maintained" relies on transcript data, but the two actually detected Lpt proteins (LptD, LptE) both decline. Acknowledge this discordance and its implications for the shared-component model.
-
Strengthen comparative analysis validation: While NB06b addresses the PaperBLAST false-negative issue, consider sequence-based homology searches (HMMER/Pfam) for the comparative claims to eliminate residual annotation-based limitations.
-
Provide appropriate caveats for single-replicate findings: The lptC2 protein induction and Pal upregulation claims rely on single-replicate data and should be presented as pilot observations requiring replication rather than established novel contributions.
Methodological Recommendations
-
Recalibrate fitness phenotype thresholds for future projects: The H2 threshold (≥10%) sitting below genome background (33.25%) represents a methodological lesson for future BERIL fitness browser analyses. Use fold-enrichment over background rather than fixed percentages.
-
Expand iron-limitation axis: The Caulobacter fitness compendium lacks iron-limitation experiments, preventing full testing of H2. Consider targeted RB-TnSeq experiments under iron chelation to complete this analysis axis.
-
Validate protein-level findings: The replicated proteomics scheduled for summer 2026 will be crucial for validating the lptC2 induction, Pal upregulation, and Lpt apparatus protein-level behavior claims.
This review was generated by an AI system. It should be treated as advisory input, not a definitive assessment.
Visualizations
00 Sphingolipid Locus Heatmap
Nb01 Fur Signature Scatter
Nb01 Leaden Iron Vs Fur
Nb04 Ctpa Per Strain
Nb04 Lpt Apparatus Heatmap
Nb04 Sphingolipid Locus Heatmap
Nb05 Pg Remodeling Heatmap
Nb06 Comparative Heatmap
Nb07 Synthesis Master
Notebooks
00_orientation.ipynb
00 Orientation
View notebook →
01_leaden2018_fur_signature.ipynb
01 Leaden2018 Fur Signature
View notebook →
02_caulo_fitness_ranking.ipynb
02 Caulo Fitness Ranking
View notebook →
02b_h2_hypergeometric_verdict.ipynb
02B H2 Hypergeometric Verdict
View notebook →
03_chvi_phase_partition_sigU.ipynb
03 Chvi Phase Partition Sigu
View notebook →
04_sphingolipid_lpt_panel.ipynb
04 Sphingolipid Lpt Panel
View notebook →
05_pg_remodeling.ipynb
05 Pg Remodeling
View notebook →
06_comparative_species.ipynb
06 Comparative Species
View notebook →
06b_ncbi_annotation_presence.ipynb
06B Ncbi Annotation Presence
View notebook →
07_synthesis.ipynb
07 Synthesis
View notebook →